Stomatal Conductance Responses of Acacia caven to Seasonal Patterns of Water Availability at Different Soil Depths in a Mediterranean Savanna


Abstract
:1. Introduction
2. Materials and Methods
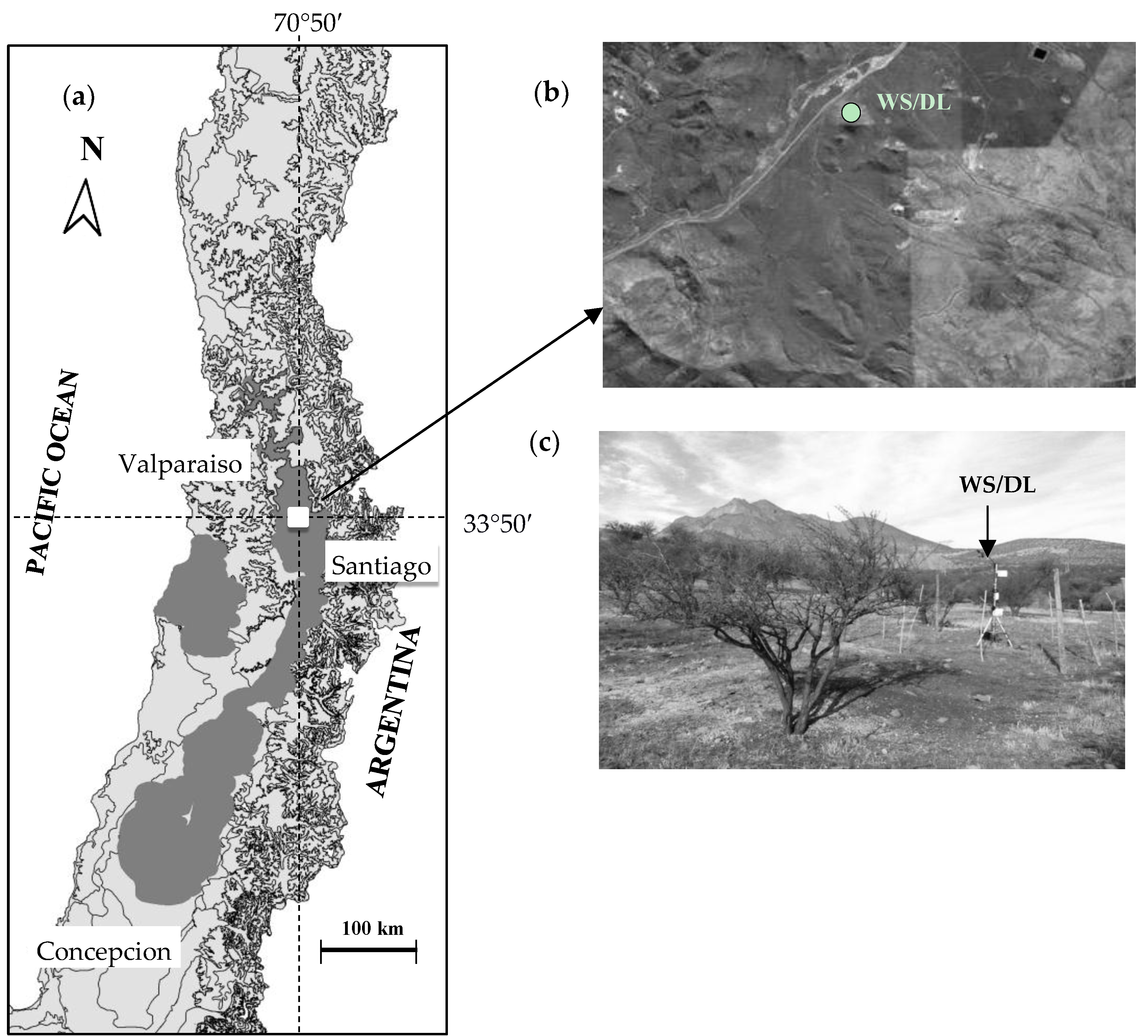
2.1. Study Area
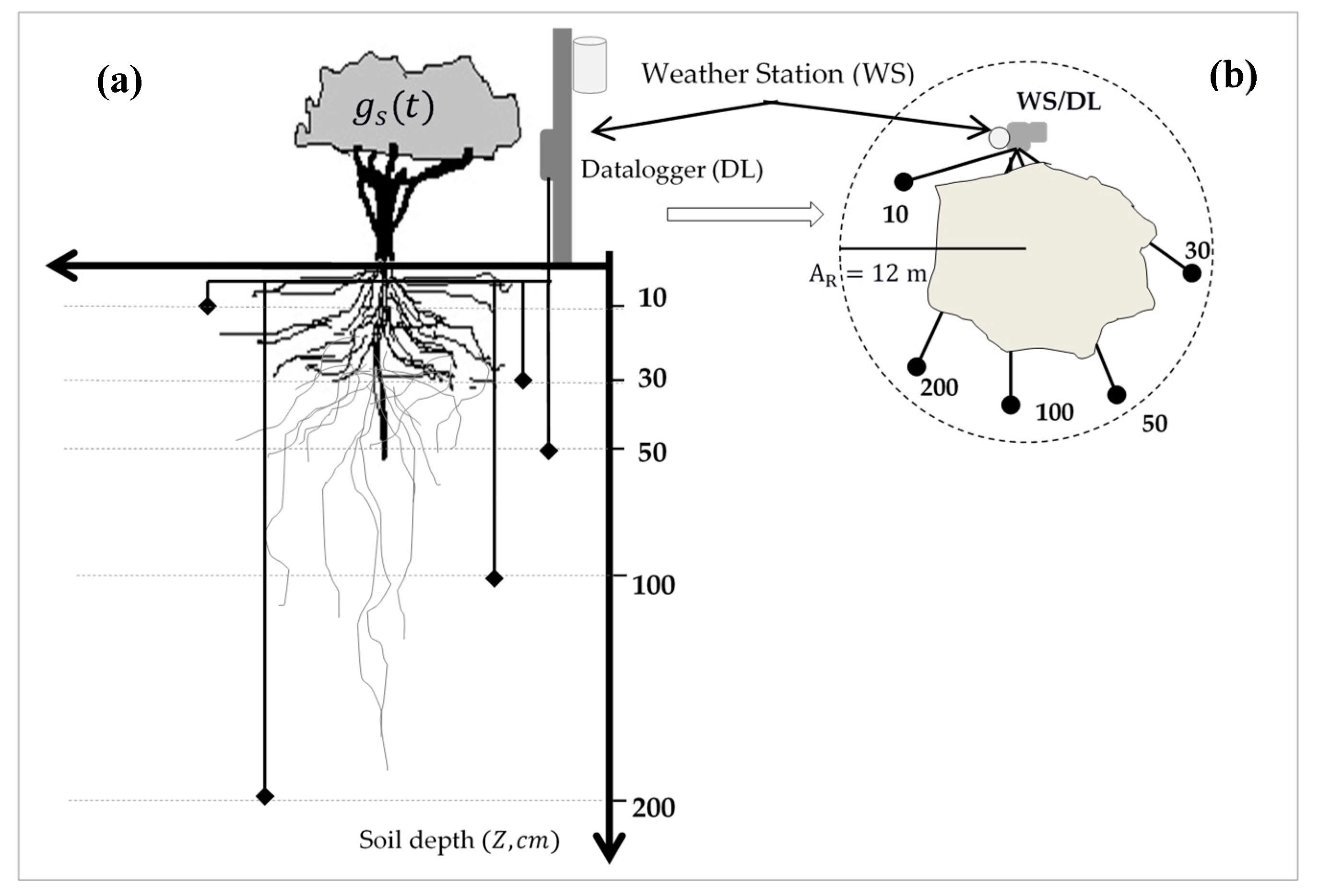
2.2. Recording the Microclimate Variables at the Site
2.3. Stomatal Conductance Measurements
2.4. Statistical Analysis
3. Results
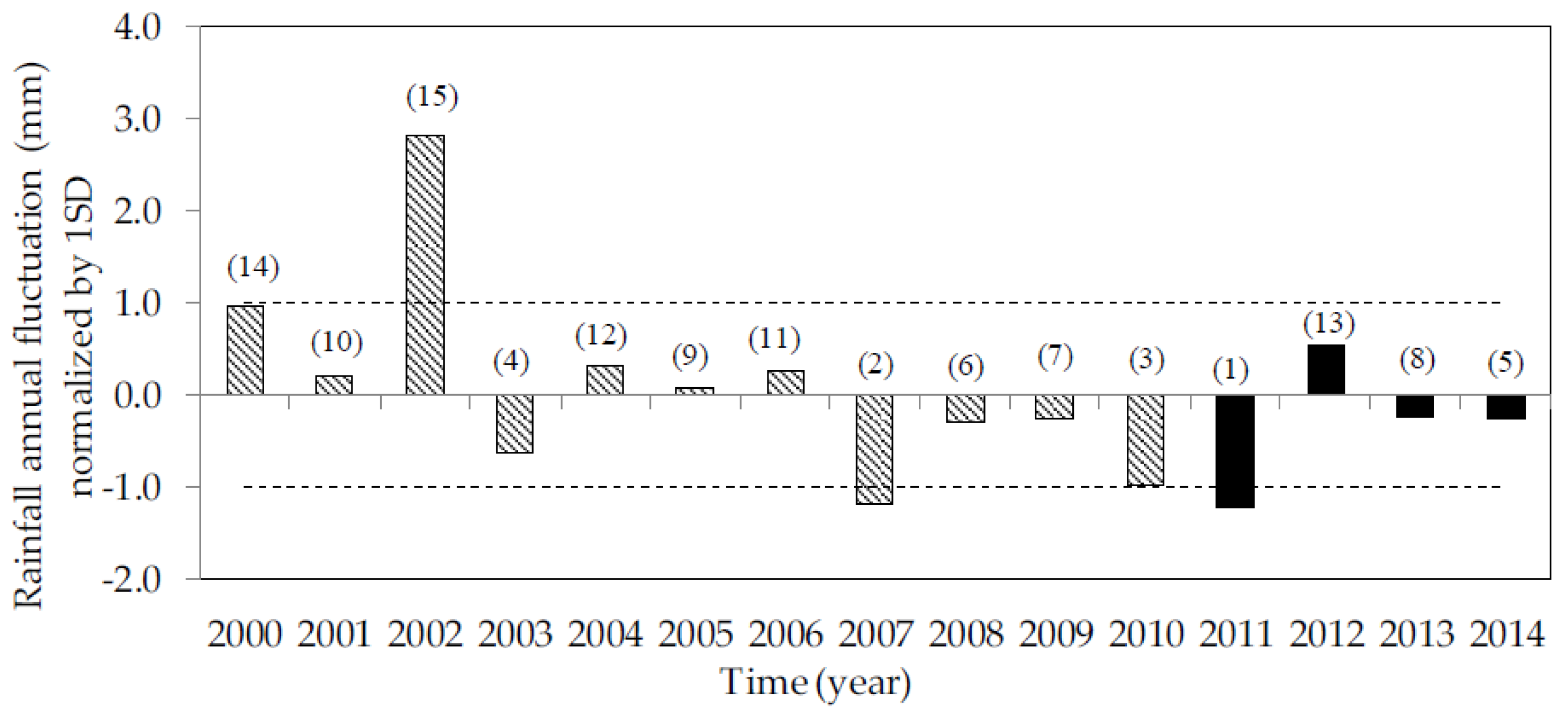
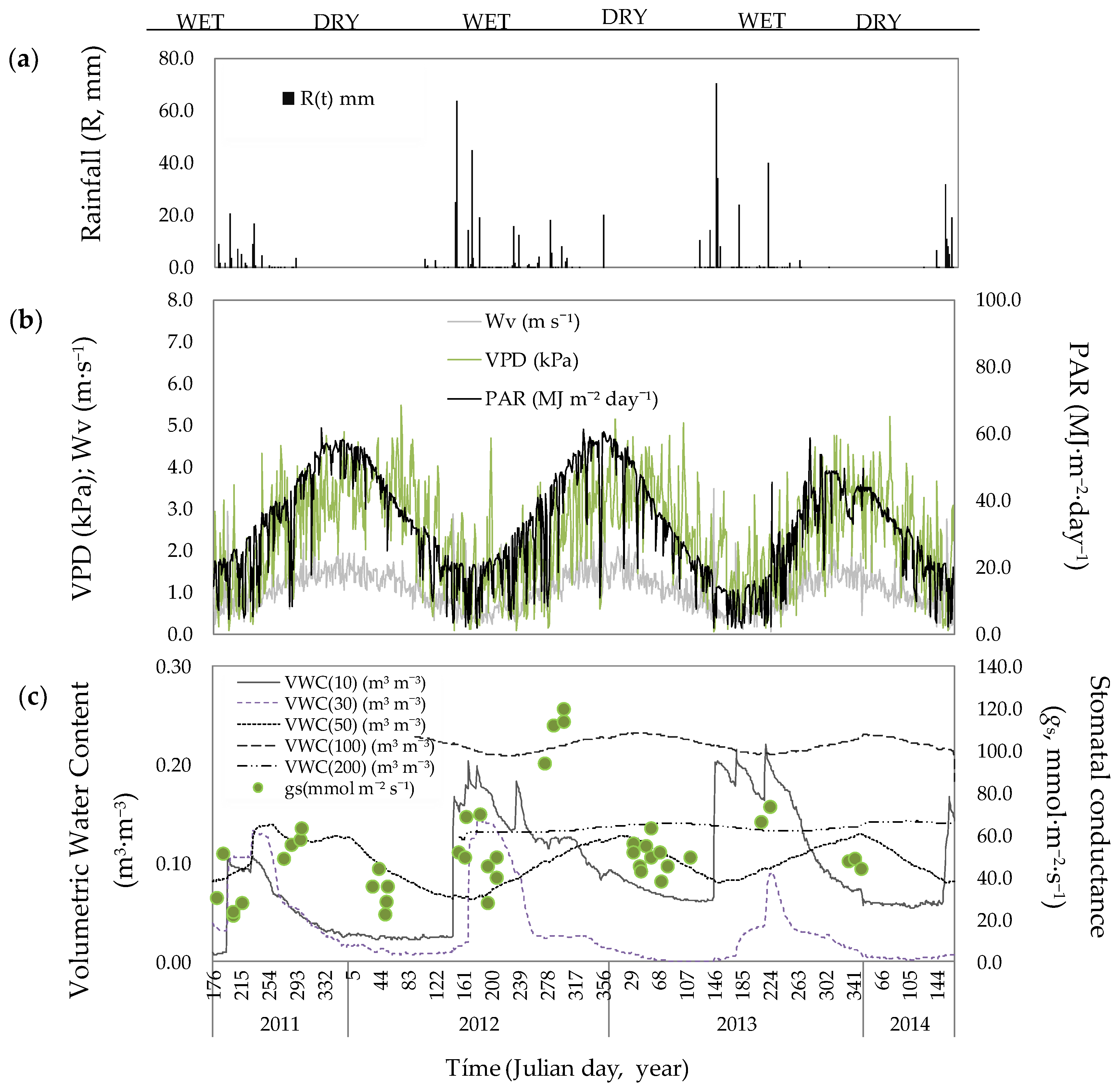
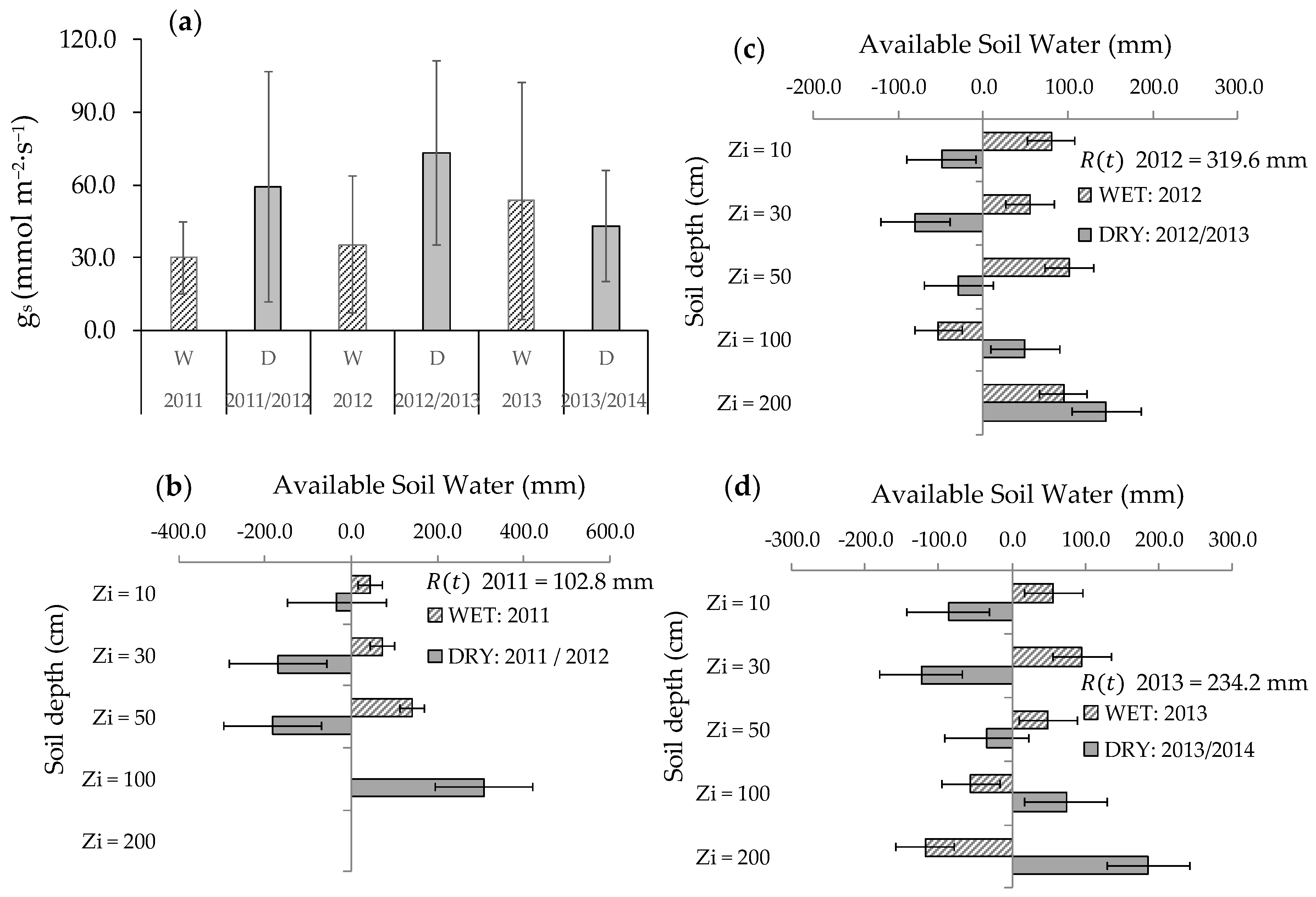
3.1. Environmental Conditions
3.2. Soil Volumetric Water Content
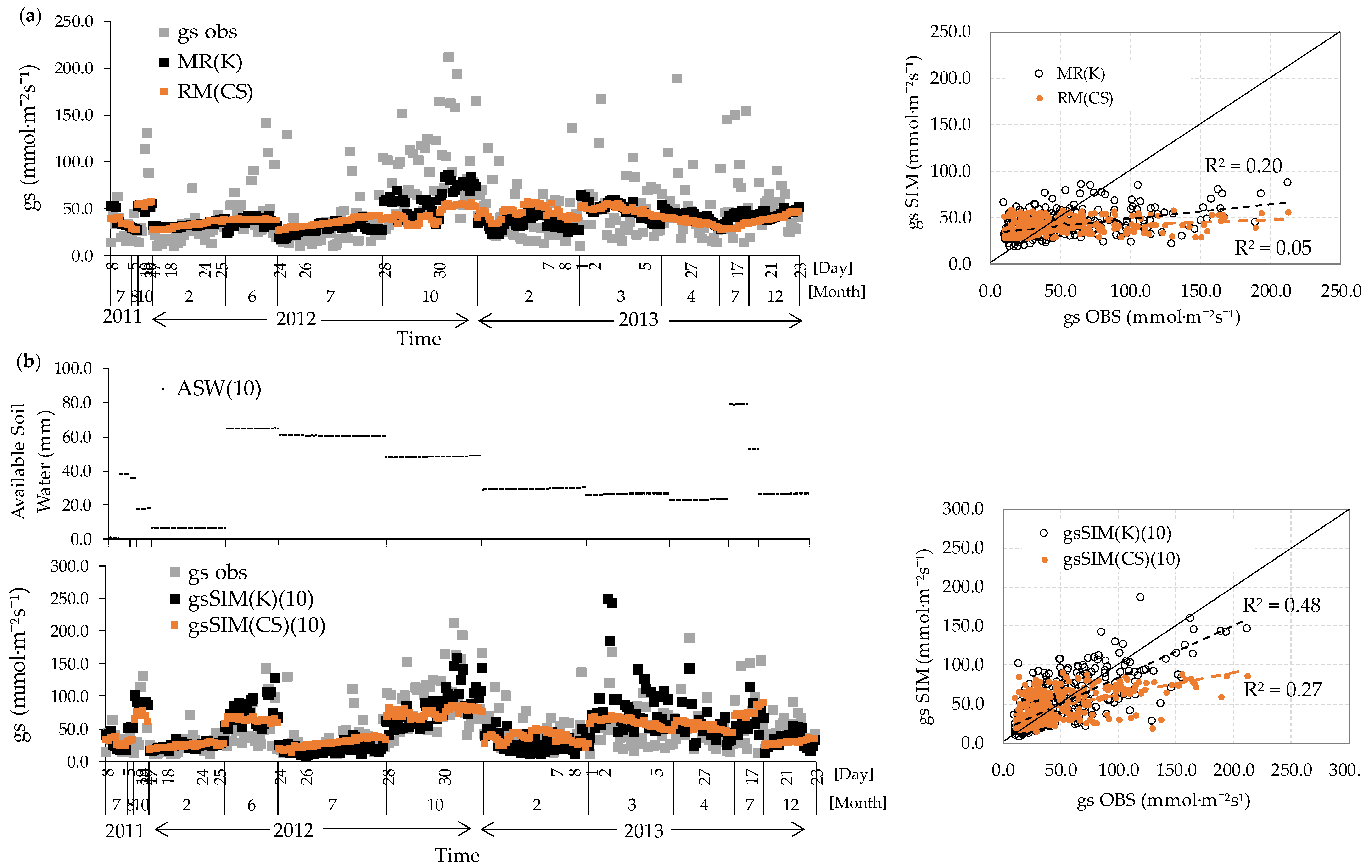
3.3. Stomatal Conductance and Vertical VWC Dynamics
3.4. Responses to Environmental Conditions
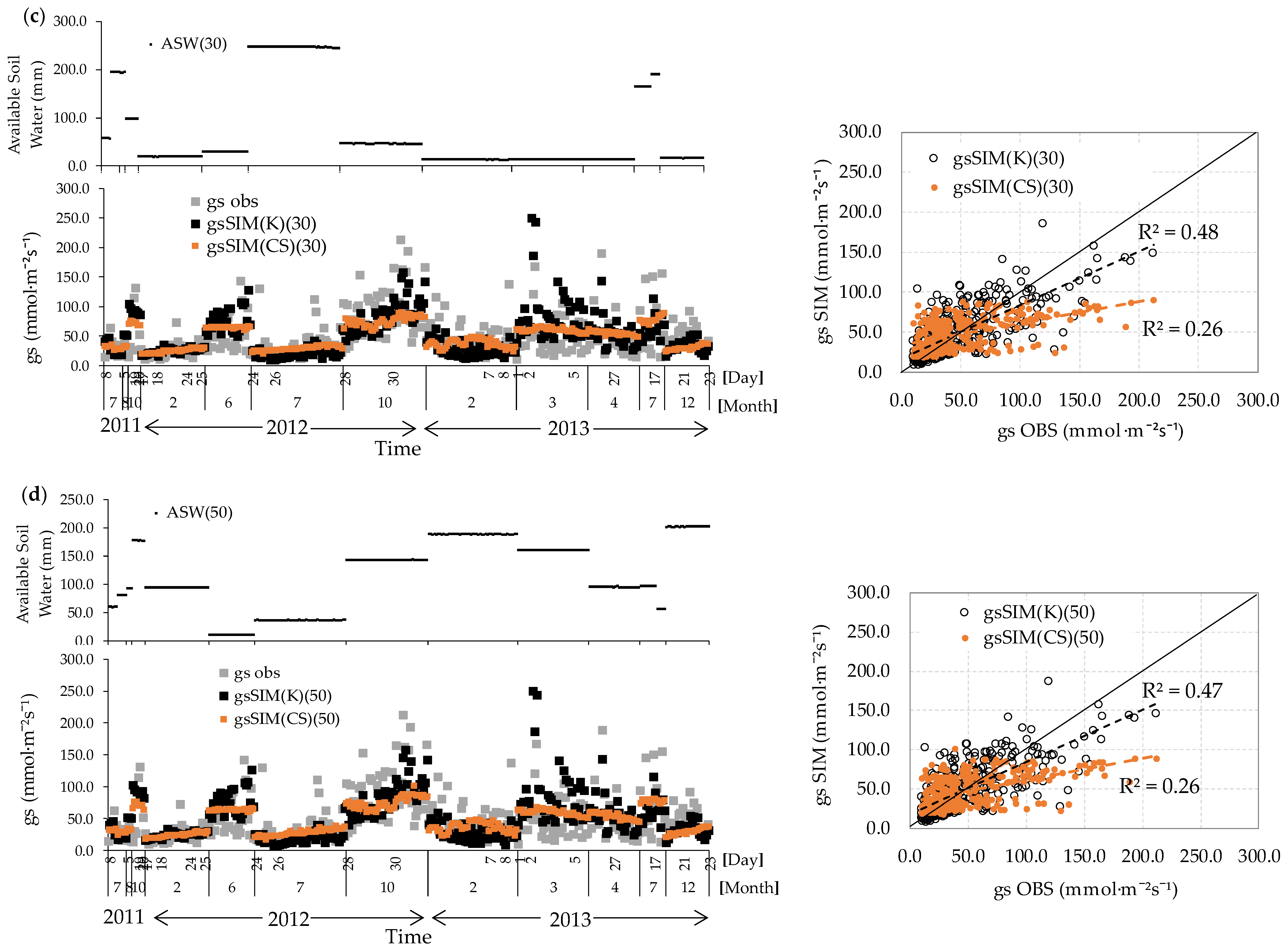
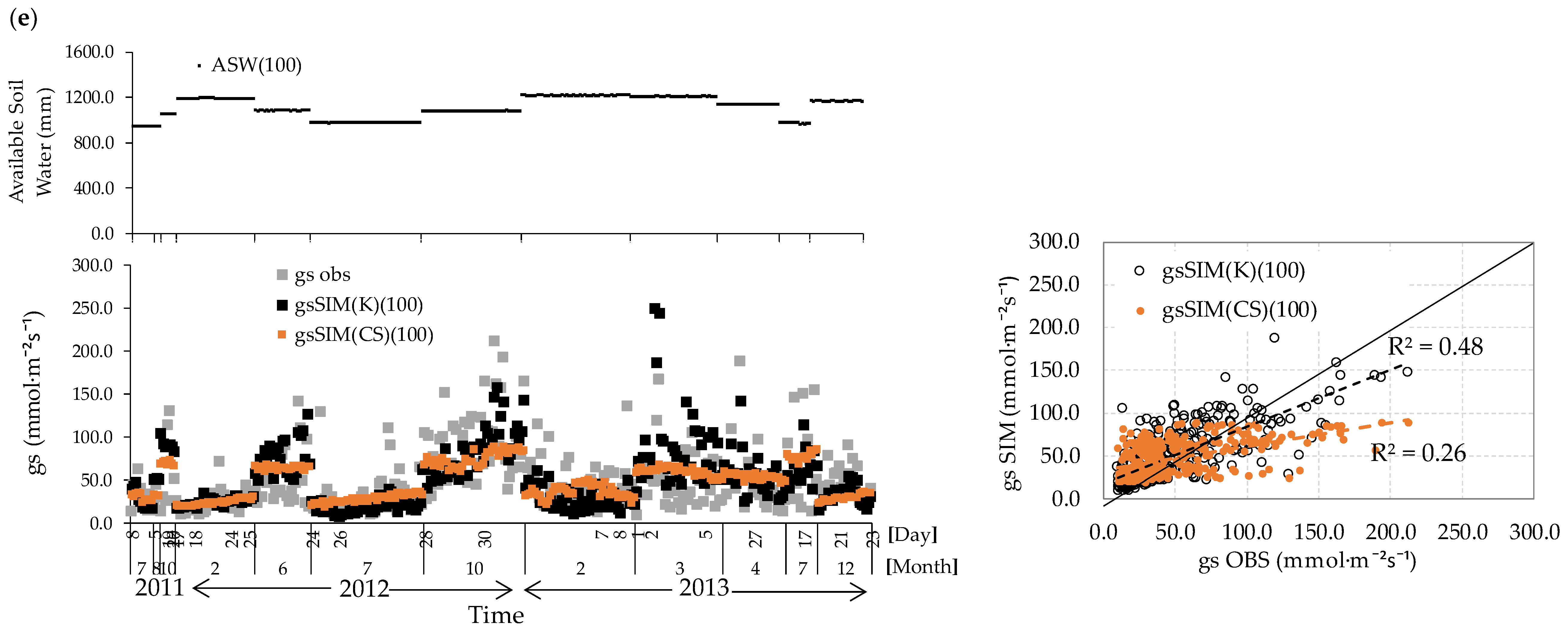
3.5. gs Responses Incorporating Volumetric Water Content
3.6. Analysis of Remaining Errors Concerning the Explanatory Variables
4. Discussion
5. Conclusions
- (1)
- A clear gain was achieved in the temporal representation of based on atmospheric variables at the level of the canopy when incorporating the VWC in the root zone.
- (2)
- The contribution of VWC at different depths in the root zone was relevant when performing an adjustment with the CS and seasonal adjustment. In the first case, VWC and not the atmospheric variables determined the seasonal fluctuation in the simulated series.
- (3)
- When we included seasonality in the adjustment of atmospheric variables, VWC was also a relevant variable.
- (4)
- No differentiated use of water within the root zone, but rather a constant activity of the radicular system, was observed as a whole in the capture of available water at 100 cm.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vico, G.; Thompson, S.E.; Manzoni, S.; Molini, A.; Albertson, J.D.; Almeida-Cortez, J.S.; Fay, P.A.; Feng, X.; Guswa, A.J.; Liu, H.; et al. Climatic, ecophysiological, and phenological control son plant ecohydrological strategies in seasonally dry ecosystems. Ecohydrology 2015, 8, 660–681. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Eco. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Tietjen, B.; Zehe, E.; Jeltsch, F. Simulating plant water availability in dry lands under climate change: A generic model of two soil layer. Water Resour. Res. 2009, 45, W01418. [Google Scholar] [CrossRef]
- Miranda, J.; Jorquera, M.J.; Pugnaire, F.I. Phenological and reproductive responses of a semiarid shrub to pulsed watering. Plant Ecol. 2014, 215, 769–777. [Google Scholar] [CrossRef]
- Katul, G.; Ellsworth, D.; Lai, C.-T. Modelling assimilation and intercellular CO2 from measured conductance: A synthesis of approaches. Plant Cell Environ. 2000, 23, 1313–1328. [Google Scholar] [CrossRef]
- Gao, Q.; Zhao, P.; Zeng, X.; Cai, X.; Shen, W. A model of stomatal conductance to quantify the relationship between leaf transpiration, microclimate and soil water stress. Plant Cell Environ. 2002, 25, 1373–1381. [Google Scholar] [CrossRef] [Green Version]
- Arve, L.E.; Torre, S.; Olsen, J.E.; Tanino, K.K. Stomatal responses to drought stress and air humidity. In Abiotic Stress in Plants—Mechanisms and Adaptations; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef]
- Schulze, E.D.; Kelliher, F.M.; Körner, C.; Lloyd, J.; Leuning, R. Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate and plant nitrogen nutrition: A global ecology scaling exercise. Annu. Rev. Ecol. Syst. 1994, 25, 629–660. [Google Scholar] [CrossRef]
- Ding, R.; Kang, S.; Du, T.; Hao, X.; Zhang, Y. Scaling up stomatal conductance from leaf to canopy using a dual-leaf model for estimating crop evapotranspiration. PLoS ONE 2014, 9, e95584. [Google Scholar] [CrossRef] [PubMed]
- Damour, G.; Simonneau, T.; Cochard, H.; Urban, L. An overview of models of stomatal conductance at the leaf level. Plant Cell Environ. 2010, 33, 1419–1438. [Google Scholar] [CrossRef] [PubMed]
- Cowan, I.R.; Farquhar, G.D. Stomatal function in relation to leaf metabolism and environment. Symp. Soc. Exp. Biol. 1977, 31, 471–505. [Google Scholar] [PubMed]
- Law, B.E.; Falge, E.; Gu, L.; Baldocchi, D.D.; Bakwin, P.; Berbigier, P.; Davis, K.; Dolman, A.J.; Falk, M.; Fuentes, J.D.; et al. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agric. For. Meteorol. 2002, 113, 97–120. [Google Scholar] [CrossRef] [Green Version]
- Schulze, E.D.; Mooney, H.A.; Sala, O.E.; Jobbagy, E.; Buchmann, N.; Bauer, G.; Canadell, J.; Jackson, R.B.; Loreti, J.; Oesterheld, M.; et al. Rooting depth, water availability, and vegetation cover along an aridity gradient in Patagonia. Oecologia 1996, 108, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Zavala, M. Integration of drought tolerance mechanisms in Mediterranean sclerophylls: A functional interpretation of leaf gas exchange simulators. Ecol. Modell. 2004, 176, 211–226. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Wolf, A.; Arango-Velez, A.; Choat, B.; Chmura, D.J.; Jansen, S.; Kolb, T.; Li, S.; Meinzer, F.; Pita, P.; et al. Plant water potential improves prediction of empirical stomatal models. PLoS ONE 2017, 12, e0185481. [Google Scholar] [CrossRef] [PubMed]
- Anav, A.; Proietti, C.; Menut, L.; Carnicelli, S.; De Marco, A.; Paoletti, E. Sensitivity of stomatal conductance to soil moisture: Implications for troposphere ozone. Atmos. Chem. Phys. 2018, 18, 5747–5763. [Google Scholar] [CrossRef]
- Oren, R.; Ewers, B.E.; Todd, P.; Phillips, N.; Katul, G. Water balance delineates the soil layer in which moisture affects canopy conductance. Ecol. Appl. 1998, 8, 990–1002. [Google Scholar] [CrossRef]
- Irvine, J.; Perks, M.P.; Magnani, F.; Grace, J. The response of Pinus sylvestris to drought: Stomatal control of transpiration and hydraulic conductance. Tree Physiol. 1998, 18, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.B. Modelling surface conductance of pine forest. Agric. For. Meteorol. 1988, 43, 19–35. [Google Scholar] [CrossRef]
- Gu, D.; Wang, Q.; Otieno, D. Canopy transpiration and stomatal responses to prolonged drought by a dominant desert species in Central Asia. Water 2017, 9, 404. [Google Scholar] [CrossRef]
- Emanuel, R.; D’Odorico, P.; Epstein, H. A dynamic soil water threshold for vegetation water stress derived from stomatal conductance models. Water Resour. Res. 2005, 43. [Google Scholar] [CrossRef]
- Granier, A.; Bréda, N.; Biron, P.; Villete, S. A lumped water balance model to evaluate duration and intensity of drought constraints in forest stands. Ecol. Modell. 1999, 116, 269–283. [Google Scholar] [CrossRef]
- Cudennec, C.; Leduc, C.; Koutsoyiannis, D. Dryland hydrology in Mediterranean regions—A review. Hydrol. Sci. J. 2007, 52, 1077–1087. [Google Scholar] [CrossRef]
- Gerstmann, C.; Miranda, M.; Condal, A. Description of space-time variability of the potential productivity of Acacia caven espinales based on MODIS images and the Enhanced Vegetation Index (EVI). Cienc. Investig. Agrar. 2010, 37, 63–73. [Google Scholar] [CrossRef]
- Ovalle, C.; Avendaño, J.; Aronson, J.; Del Pozo, A. Land occupation patterns and vegetation structure in the anthropogenic savannas (espinales) of central Chile. For. Ecol. Manag. 1996, 86, 129–139. [Google Scholar] [CrossRef]
- Aronson, J.; Ovalle, C.; Aguilera, L.; Leon, P. Phenology of an ‘inmigrant’ savanna tree (Acacia caven, Legumoniosae) in the Mediterranean climate zone of Chile. J. Arid Environ. 1994, 27, 55–70. [Google Scholar] [CrossRef]
- Schultz, J.J.; Cayuela, L.; Rey-Benayas, J.M.; Schröder, B. Factors influencing vegetation cover change in Mediterranean Central Chile (1975–2008). Appl. Veg. Sci. 2011, 14, 571–582. [Google Scholar] [CrossRef]
- Van de Wouw, P.; Echeverria, C.; Rey-Benayas, J.M.; Holmgren, M. Persistent Acacia savannas replace Mediterranean sclerophyllous forests in South America. For. Ecol. Manag. 2011, 262, 1100–1108. [Google Scholar] [CrossRef]
- Raab, N.; Meza, F.J.; Franck, N.; Bambach, N. Empirical stomatal conductance models revel that the isohydric behavior of an Acacia caven Mediterranean Savannah scales from leaf to ecosystem. Agric. For. Meteorol. 2013, 213, 203–216. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Sepulveda, M.; Bown, E.H.; Miranda, M.D.; Fernandez, B. Impact of rainfall frequency and intensity on inter- and intra-annual satellite-derived EVI vegetation productivity of an Acacia caven shrubland community in Central Chile. Plant Ecol. 2018. [Google Scholar] [CrossRef]
- Allen, R.G.; Pruitt, W.O.; Raes, D.; Smith, M.; Pereira, L.S. Estimating evaporation from base soil and the crop coefficient for the initial period using common soils information. J. Irrig. Drain. Eng. 2005, 131, 14–23. [Google Scholar] [CrossRef]
- Abtew, W.; Melesse, A.M. Evaporation and Evapotranspiration: Measurements and Estimations; Springer: Argovie, The Netherlands, 2013; ISBN 978-94-007-4737-1. [Google Scholar]
- Blake, G.R. Bulk density. In Methods of Soil Analysis: Part 1, Physical and Mineralogical Properties, Including Statistics of Measurement and Sampling; Black, C.A., Evans, D.D., White, J.L., Ensminger, L.E., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 374–390. [Google Scholar]
- Johnson, M.-V.; Kiniry, J.; Burson, B. Ceptometer deployment method affects measurement of fraction of intercepted photosynthetically active radiation. Agron. J. 2010, 102, 1132–1137. [Google Scholar] [CrossRef]
- Zuur, A.F.; Leno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Meth. Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Matsumoto, K.; Ohta, T.; Tanaka, T. Dependence of stomatal conductance on leaf chlorophyll concentration and meteorological variables. Agric. For. Meteorol. 2005, 132, 44–57. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoids x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.L.; Niemann, J.D. An evaluation of nonlinear methods for estimating catchment-scale soil moisture patterns based on topographic attributes. J. Hydroinf. 2012, 14, 800–814. [Google Scholar] [CrossRef] [Green Version]
- Wooldridge, J.M. Introductory Econometrics: A Modern Approach, 5th International ed.; South-Western: Mason, OH, USA, 2013; pp. 409–415. [Google Scholar]
- Bennett, N.D.; Croke, G.F.W.; Guariso, G.; Guillaume, J.H.A.; Hamilton, S.H.; Jakeman, A.J.; Marsili-Libelli, S.; Newham, L.T.H.; Norton, J.P.; Perrin, C.; et al. Characterizing performance of environmental models. Environ. Modell. Softw. 2013, 40, 1–20. [Google Scholar] [CrossRef]
- Legates, D.R.; McCabe, G.J. Evaluating the use of “goodness-of-fit” measures in hydrologic and hydroclimatic model validation. Water Resour. Res. 1999, 35, 233–241. [Google Scholar] [CrossRef]
- Harris, P.P.; Huntingford, C.; Cox, P.M.; Gash, J.H.C.; Malhi, Y. Effect of soil moisture on canopy conductance of Amazonian rainforest. Agric. For. Meteorol. 2004, 122, 215–227. [Google Scholar] [CrossRef]
- Santra, P.; Kumar, M.; Kumawat, R.N.; Painuli, D.K.; Hati, K.M.; Heuvelink, G.B.M.; Batjes, N.H. Pedotransfer functions to estimate soil water content at field capacity and permanent wilting point in hot Arid Western India. J. Earth Syst. Sci. 2018, 127, 35. [Google Scholar] [CrossRef]
- Meza, F.J.; Montes, C.; Bravo-Martinez, F.; Serrano-Ortiz, P.; Kowalski, A.S. Soil water content effects on net ecosystem CO2 exchange and actual evapotranspiration in a Mediterranean semiarid savanna of Central Chile. Sci. Rep. 2018, 8, 8570. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Gan, Y.; Chen, P.; Qiu, M.; Jiang, K.; Wang, G. Stomata prioritize their responses to multiple biotic and abiotic signal inputs. PLoS ONE 2014, 9, e101587. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, X.; Zhang, Z.; He, C.; Zhao, P.; You, Y.; Mo, L. Seasonal changes in Cyclobalanopsis glauca transpiration and canopy stomatal conductance and their dependence on subterranean water and climatic factors in rocky karst terrain. J. Hydrol. 2011, 402, 135–143. [Google Scholar] [CrossRef]
- Williams, M.; Malhi, Y.; Nobre, A.D.; Rastetter, E.B.; Grace, J.; Pereira, M.G.P. Seasonal variation in net carbon exchange and evapotranspiration in a Brazillian rain forest: A modelling analysis. Plant Cell Environ. 1998, 21, 953–968. [Google Scholar] [CrossRef]
- Oren, R.; Pataki, D.E. Transpiration in response to variation in microclimate and soil moisture in southeastern deciduous forests. Oecologia 2001, 127, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Baldocchi, D.D. Seasonal trends in photosynthetic parameters and stomatal conductance of blue oak (Quercus douglasii) under prolonged summer drought and high temperature. Tree Physiol. 2003, 23, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Mediavilla, S.; Escudero, A. Stomatal responses to drought of mature trees and seedlings of two co-ocurring Mediterranean oaks. For. Ecol. Manag. 2004, 187, 281–294. [Google Scholar] [CrossRef]
- Uddling, J.; Teclaw, R.M.; Pregitzer, K.S.; Ellsworth, D.S. Leaf and canopy conductance in aspen and aspen-birch forests under free-air enrichment of carbon dioxide and ozone. Tree Physiol. 2009, 29, 1367–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman, D.T.; Novick, K.A.; Brzostek, E.R.; Dragoni, D.; Rahman, F.; Phillips, R.P. The role of isohydric and anisohydric species in determining ecosystem-scale to severe drought. Oecologia 2015, 179, 641–654. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Un | Soil Layer Depth (i) in cm | ||||
|---|---|---|---|---|---|
| i = 10 | i = 30 | i = 50 | i = 100 | ||
| Field capacity | % weight | 40.8 ± 4.8 | 40.2 ± 4.3 | 41.6 ± 3.9 | 47.1 ± 4.5 |
| Wilting point | % weight | 24.4 ± 3.2 | 25.5 ± 3.5 | 26.6 ± 4.2 | 30.6 ± 4.6 |
| Organic Matter | % | 1.8 ± 1.2 | 2.5 ± 1.4 | 3.9 ± 2.3 | 2.5 ± 1.6 |
| Bulk density | g·cm−3 | 1.19 ± 0.08 | 1.16 ± 0.06 | 1.08 ± 0.06 | 1.13 ± 0.05 |
| Texture | Sand (%) | 21 ± 6.4 | 10 ± 3.2 | ||
| Silt (%) | 28 ± 4.2 | 20 ± 3.1 | |||
| Clay (%) | 51 ± 8.7 | 70 ± 5.1 | |||
| Total porosity | % | 55 ± 5.3 | 51 ± 5.5 | 51 ± 4.5 | 50 ± 4.5 |
| (p < 0.05) | |||
|---|---|---|---|
| Variables | CS | D(t) | W(t) |
| 0.013 | −0.364 | 0.319 | |
| 0.070 | 0.141 | 0.004 | |
| PAR | 0.231 | 0.389 | −0.101 |
| VPD | 0.026 | 0.001 | 0.132 |
| Intercept | 3.27 | 4.57 | 2.63 |
| n | 329 | 227 | 102 |
| D-W | 1.15 | 1.07 | 1.11 |
| (%) | 5.4 | 17.8 | 12.3 |
| 19.5 | |||
| MCE | 0.10 | 0.16 | |
| MAE | 24.55 | 23.04 | |
| (a) | ||||||||
| 10 | 30 | 50 | 100 | 10 | 30 | 50 | 100 | |
| 0.002 | 0.005 | 0.005 | 0.003 | 0.002 | 0.005 | 0.005 | 0.003 | |
| −0.029 | −0.028 | −0.021 | −0.027 | −0.029 | −0.028 | −0.021 | −0.027 | |
| 0.038 | −0.017 | −0.001 | 0.029 | 0.038 | −0.017 | −0.001 | 0.029 | |
| −0.017 | 0.049 | 0.096 | −0.027 | −0.017 | 0.049 | 0.096 | −0.027 | |
| (b) | ||||||||
| 10 | 30 | 50 | 100 | 10 | 30 | 50 | 100 | |
| MCE | 0.49 | 0.48 | 0.49 | 0.49 | 0.44 | 0.44 | 0.45 | 0.44 |
| MAE | 13.96 | 14.27 | 14.05 | 14.16 | 15.33 | 15.36 | 15.22 | 15.34 |
| R2 | 47.7% | 47.9% | 47.2% | 47.6% | 26.8% | 26.2% | 25.6% | 26.0% |
| MR | DR Incorporating | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Seasonal (K) | Complete Serie (CS) | |||||||||
| µ(K) | µ(CS) | µ(10) | µ(30) | µ(50) | µ(100) | µ(10) | µ(30) | µ(50) | µ(100) | |
| Av | 1.21 | 1.24 | 1.10 | 1.10 | 1.11 | 1.10 | 1.09 | 1.08 | 1.09 | 1.08 |
| SD | 0.82 | 0.89 | 0.54 | 0.54 | 0.56 | 0.54 | 0.71 | 0.68 | 0.70 | 0.69 |
| CV | 0.68 | 0.72 | 0.49 | 0.49 | 0.51 | 0.49 | 0.65 | 0.63 | 0.65 | 0.64 |
| Max | 6.14 | 5.22 | 4.62 | 4.60 | 4.89 | 4.76 | 6.92 | 5.27 | 5.78 | 5.49 |
| Min | 0.15 | 0.19 | 0.13 | 0.13 | 0.13 | 0.13 | 0.16 | 0.16 | 0.16 | 0.17 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepulveda M., M.; Bown, H.E.; Fernandez L., B. Stomatal Conductance Responses of Acacia caven to Seasonal Patterns of Water Availability at Different Soil Depths in a Mediterranean Savanna. Water 2018, 10, 1534. https://doi.org/10.3390/w10111534
Sepulveda M. M, Bown HE, Fernandez L. B. Stomatal Conductance Responses of Acacia caven to Seasonal Patterns of Water Availability at Different Soil Depths in a Mediterranean Savanna. Water. 2018; 10(11):1534. https://doi.org/10.3390/w10111534
Chicago/Turabian StyleSepulveda M., Marcelo, Horacio E. Bown, and Bonifacio Fernandez L. 2018. "Stomatal Conductance Responses of Acacia caven to Seasonal Patterns of Water Availability at Different Soil Depths in a Mediterranean Savanna" Water 10, no. 11: 1534. https://doi.org/10.3390/w10111534