Soil Layer Development and Biota in Bioretention

1
Designgreen, LLC, 524 Ava Way NE, Washington, DC 20017, USA
2
Department of Environmental Science and Technology, University of Maryland, College Park, MD 20742, USA
*
Author to whom correspondence should be addressed.
Water 2018, 10(11), 1587; https://doi.org/10.3390/w10111587
Submission received: 5 September 2018
/
Revised: 28 October 2018
/
Accepted: 2 November 2018
/
Published: 6 November 2018
(This article belongs to the Special Issue Plant and Microbial Processes in Stormwater Treatment Systems)
Abstract
:As bioretention comes into widespread use, it has become increasingly important to understand the development of bioretention soils over time. A field survey was conducted to investigate the development of bioretention soils and soil ecosystems. Sampling from 10 bioretention cells of various ages provides the first detailed descriptions of bioretention soil profiles. The study reveals much biological activity in bioretention and evidence of pedogenesis even in very young sites. The uppermost soil layers were found to be enriched with organic matter, plant roots, and soil organisms. This survey provides a first glimpse into the biological processes at work in bioretention soils. The research shows that soil organisms are ubiquitous in bioretention cells and suggests that their impact on bioretention performance may be significant.
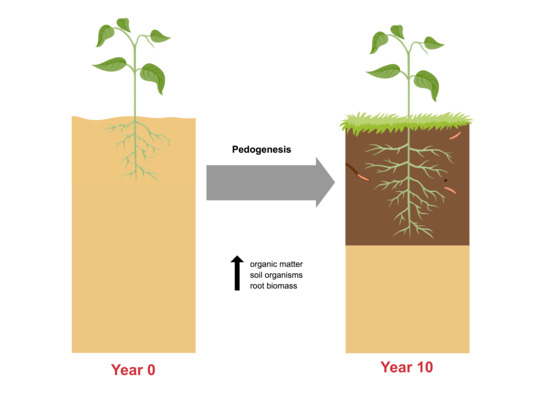
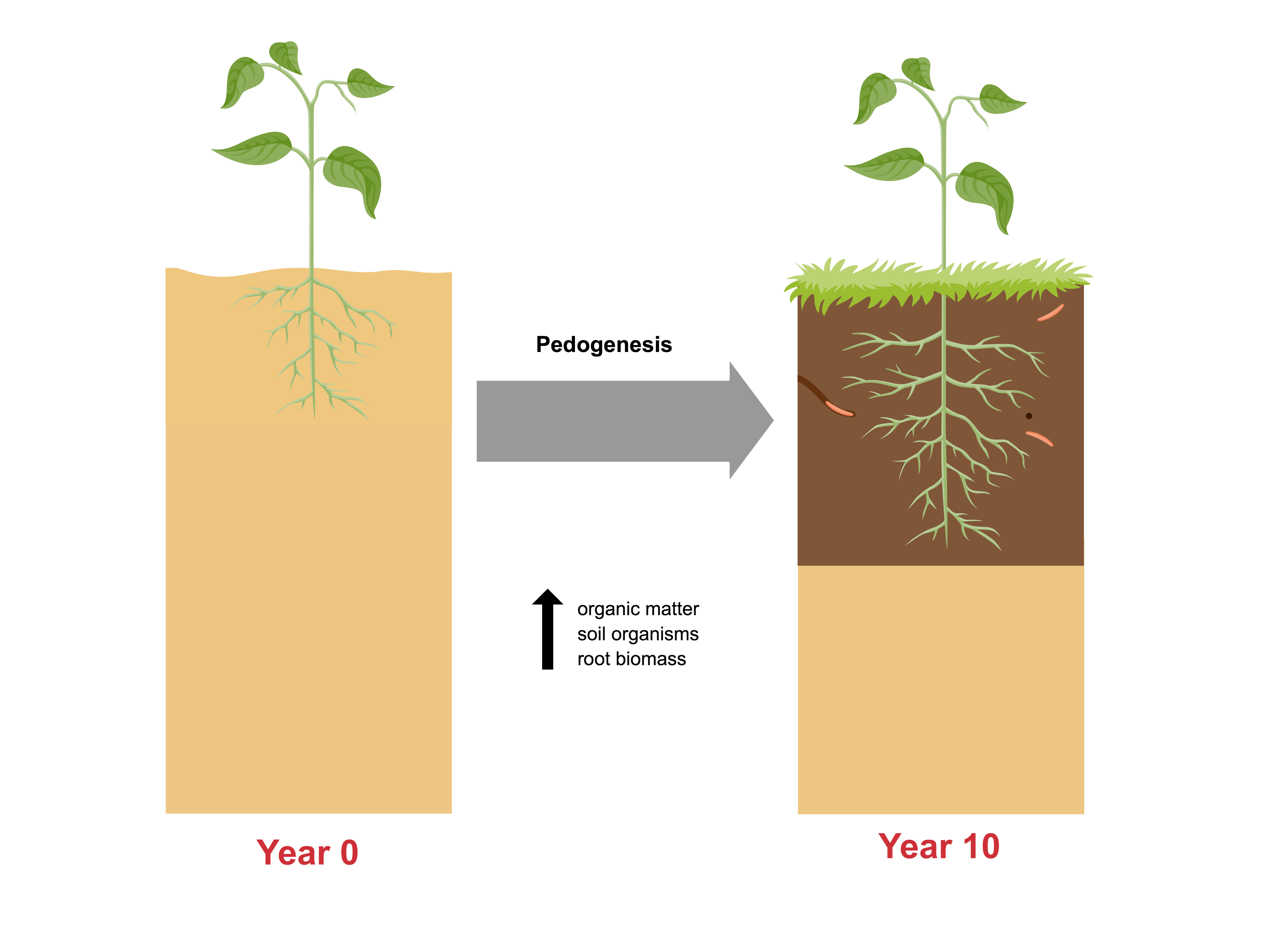
1. Introduction
Bioretention cells are a stormwater best management practice (BMP) that are commonly used in low-impact development (LID) and green infrastructure designs. The increasing popularity of bioretention cells as stormwater control structures highlights the need for a more thorough understanding of bioretention performance. Bioretention cells are essentially planted soil filters, and their performance during a storm event depends on the hydraulic conductivity and pollutant removal capacity of the soil medium. These soil properties are influenced by the physical effects of storm events as well as the actions of the plants, microbes, and soil animals of the bioretention ecosystem. Soil scientists use the term “pedogenesis” to describe the gradual changes in a soil that occur naturally over time in response to weathering, plant growth, and decomposition, and the activities of soil animals such as earthworms [1]. The bioretention soil medium can be expected to undergo pedogenesis as every soil does, beginning with the development of organically and biologically enriched horizons at the soil surface [2]. As this topsoil develops, it may change the soil media’s physical and chemical properties, and therefore affect performance. Thus, the development of bioretention soils over time must be examined if the long-term performance of bioretention is to be understood.
The potential for the accumulation of fines in bioretention cells over time and the potential for this to negatively impact hydraulic conductivity are a subject of interest to bioretention researchers. Bioretention cells intercept and filter stormwater runoff which often contains fines and would therefore be expected to experience an increase in fines over time. Li and Davis have studied the potential for bioretention systems to become clogged with sediment and found evidence of sediment accumulation in the upper soil layers [3,4]. Emerson and Traver [5] conducted a four-year study of the hydrologic performance of a bioretention cell, finding seasonal variations in infiltration rate, but no evidence of clogging over time. Jenkins, Wadzuk, and Welker [6] observed the accumulation of fines in a bioretention cell over several years but did not observe a corresponding change in infiltration rate. Le Coustumer et al. [7], however, found that the hydraulic conductivity of bioretention cells did tend to decrease over time. In their measurements of hydraulic conductivity in 37 bioretention cells, they found that hydraulic conductivity decreased to an average of half its initial value in cells up to three years old.
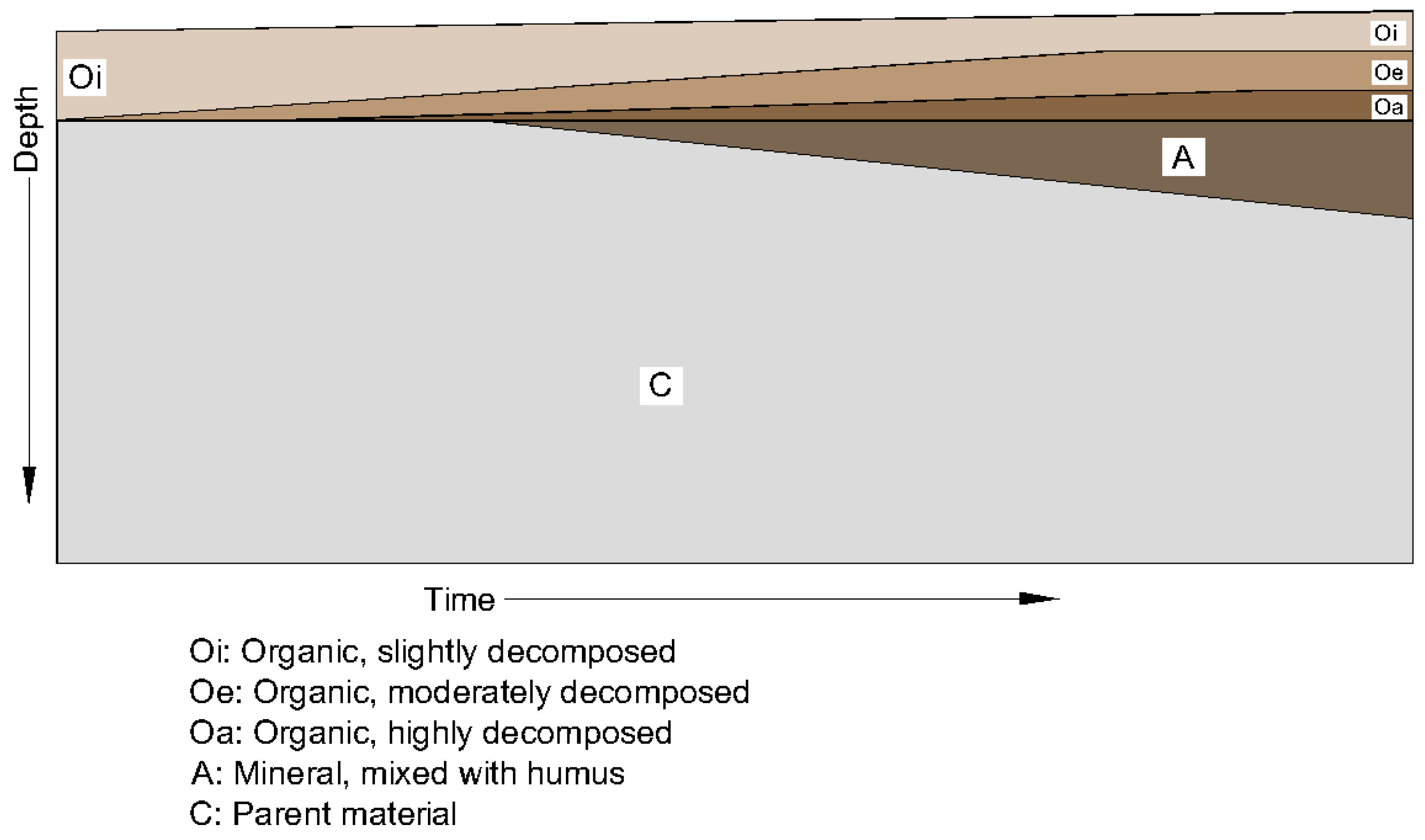
Researchers interested in the long-term behavior of bioretention soils can look to the soil science literature for clues as to how these soils might change over time in response to sediment deposition and colonization by plants and animals. The development of newly established soils tends to follow a characteristic pattern, in which the soil surface is colonized by vegetation, which adds organic matter to the soil surface through litter deposition, root exudates, and root growth and dieback [8]. Over the first few years, an organic (O) horizon forms at the soil surface, followed by a gradually thickening A horizon just below the soil surface, as illustrated in Figure 1. The A horizon, commonly referred to as “topsoil”, is distinguished by its darker color, resulting from enrichment if the parent material (C) with organic material from the O horizon.
Even at the initial stages, pedogenetic changes occur over the course of years, so a common study method is to use a space-for-time substitution technique known as a chronosequence [9]. In a chronosequence, a set of research sites are selected between which the only major variable is time. Chronosequences have, for example, been used to study the recovery of ecosystems in areas disturbed by mining [10,11,12]. Abandoned open-pit mines and piles of mine spoils are similar to bioretention cells in that they can be viewed as disturbed soils where the clock of soil and ecological development has been effectively reset, presenting valuable opportunities to observe primary succession and the early stages of pedogenesis. Leisman [10] studied primary succession on spoil banks from iron mining in Minnesota using a chronosequence. The depth of the A horizon increased monotonically from zero to 1 inch (2.54 cm) over the first 20 years. Organic matter content started out very low, and gradually increased over time, from 0.08% at 2 years to 1.26% at 51 years at the 1-inch (2.54 cm) depth. Similar increases in organic matter in the uppermost soil layers have been observed at a wide range of sites [10,12,13,14,15].
These early increases in soil organic matter are controlled by ecological development both within the soil and at its surface. The establishment of a plant community is accompanied by colonization of soil- and surface-dwelling macroinvertebrates, who carry out essential material cycling functions, facilitating the decomposition of plant material and its incorporation into the soil matrix [16,17]. Earthworms play an essential and pivotal role in these processes [18].
This research cataloged variations between bioretention soils of different ages, with an emphasis on soil ecosystem development. These data yielded a chronosequence showing the evolution of biological activity and soil profiles over time. While comparison between the research sites was complicated by differences in design and history, these data provided a valuable glimpse into processes working at a much larger time scale than are practical to study experimentally. Macroinvertebrate density and species richness are expected to increase over time during the early development of the bioretention ecosystem. Soil organic matter is expected to increase in the upper soil layers as the bioretention ecosystem begins to establish first a litter layer and then an organically-enriched topsoil layer as soil animals incorporate plant detritus into the soil.
2. Materials and Methods
Surveys were conducted of 10 existing bioretention cells of various ages. The purpose of these surveys was to assess the level of biological activity in the bioretention cells and to characterize their soil profiles. The data collected included earthworm quantity, species, and size, soil organic matter, soil particle size distribution, root biomass, macroinvertebrate abundance, and species richness.
2.1. Field Sites
Field sites were selected to represent a wide range of bioretention ages and design styles, in order to understand the full spectrum of bioretention in use in the region. Bioretention cells ranged in age from 1 year to 10 years at the time of sampling. Of the 10 field sites, 9 are located in the State of Maryland. One site, Washington Navy Yard, is located in Washington, DC. Field sites are identified and described in Table 1. Sampling was conducted during the summers of 2004 and 2005.
Specifications for the bioretention soil medium vary by municipality and have changed over time. In 2000, the State of Maryland published a Stormwater Design Manual, establishing a statewide standard for bioretention media [19] (p. B.4.7). Bioretention cells constructed prior to 2000 were not subject to this standard. Where construction documents were available, the bioretention media specification is included in Table 1. Unfortunately, as-built media testing results were not available for any of the field sites.
2.2. Field Sampling
Each site was sampled at three locations spaced evenly along a transect spanning the length of the bioretention. At each location, the litter layer was removed and plants were clipped at the soil surface. A 20 × 30 cm hole was dug, and the first 10 cm of soil was removed. This material was hand sorted. Plant roots were collected. Earthworm lengths were recorded. Representatives of different earthworm species were then sedated in a weak ethanol solution and preserved in a 4% formalin solution for later identification. Other macroscopic invertebrates were tallied and returned to the bioretention. Macroinvertebrates found on the soil surface or within the soil (henceforth referred to as “soil animals”) were classified into easily identifiable taxa. The volume of the removed soil was recorded. Approximately 1 L of soil was removed for laboratory analysis. The procedure was repeated for the soil from 10–20 cm depth, and from 20–30 cm depth.
2.3. Laboratory Analysis
Plant roots were washed, and live roots were separated from dead. Live roots were characterized by flexibility, light-colored interior, showing no signs of decay. Roots considered dead were dark throughout, brittle, or decayed. The live roots were dried overnight in a 70 °C oven, and then weighed.
Soil organic matter (SOM) was measured using the loss-on-ignition method [20]. A hydrometer was used to quantify the proportions of sand, silt, and clay in the samples. Samples with more than 10% organic matter were first oxidized using a 6% hydrogen peroxide solution. The resulting particle size distribution curve was used to calculate the coefficient of uniformity (d60/d10), where d60 and d10 are the soil particle diameters for which 60% and 10% of the mass of a soil sample is finer, respectively.
A taxonomic guide to earthworms commonly found in Maryland, as catalogued by Csuzdi and Slavecz [21] and Reynolds [22], was developed by compiling descriptions from several sources [23,24,25,26,27,28,29,30,31,32,33,34,35,36,37]. The preserved earthworm samples were examined using a dissecting microscope. Earthworms were identified as completely as possible by external features using the key. The extent to which the earthworm samples were identifiable depended upon their maturity, and on the presence or absence of unique identifying features. Non-clitellate (immature) specimens lack many identifying features, and can rarely be identified, even to genus.
3. Results
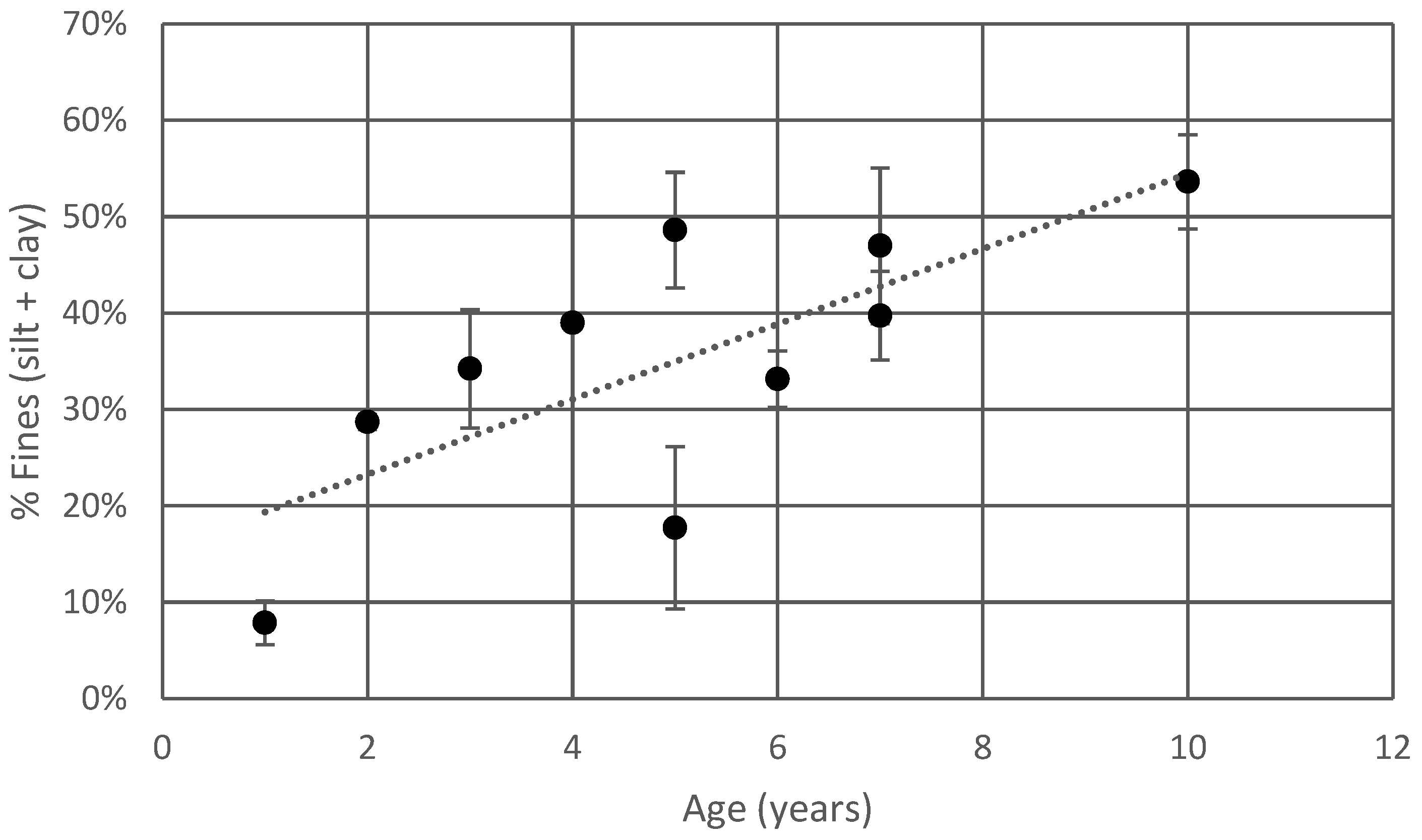
Particle size analysis data are presented in Table 2. Fines (combined silt and clay fractions) were observed to increase with the age of the bioretention cell (Figure 2).
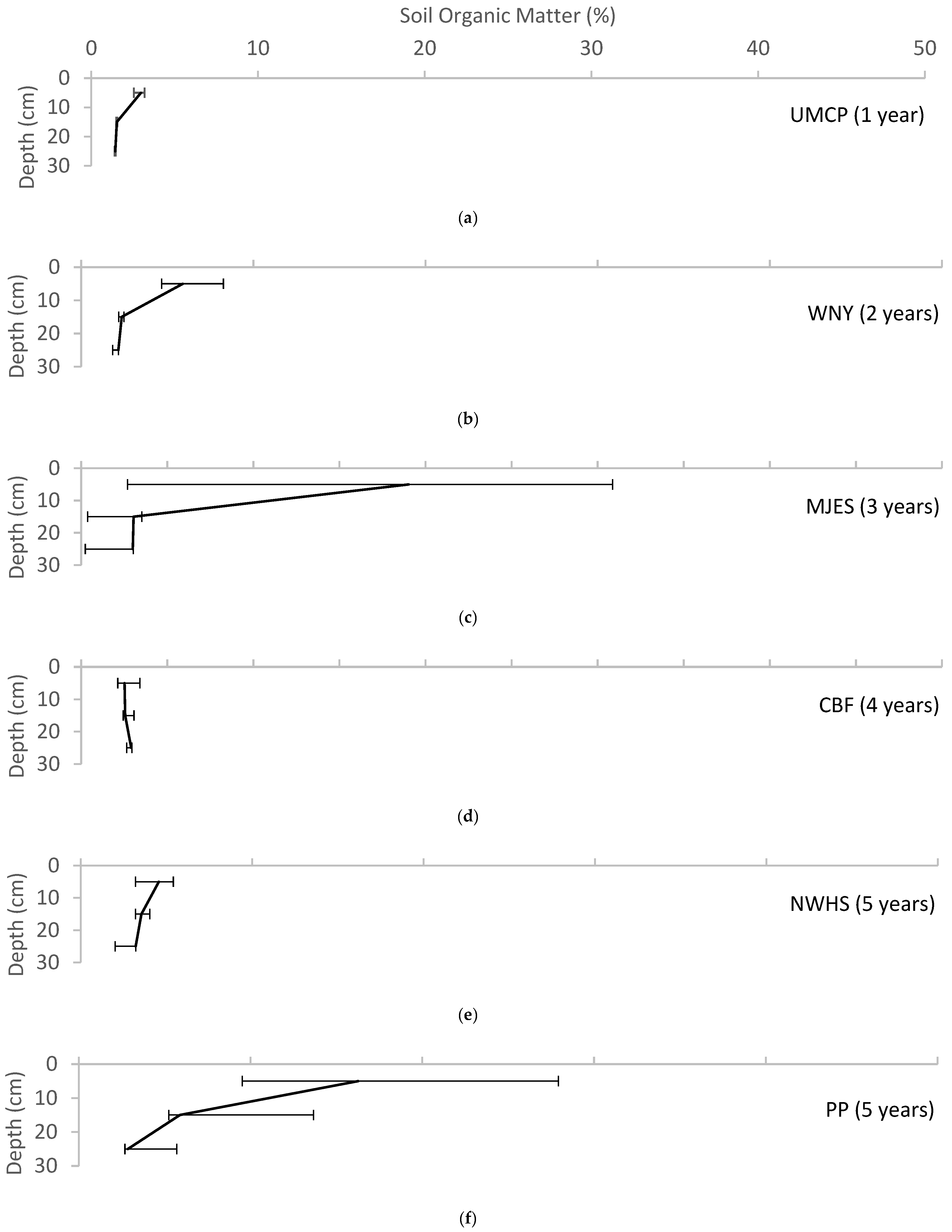
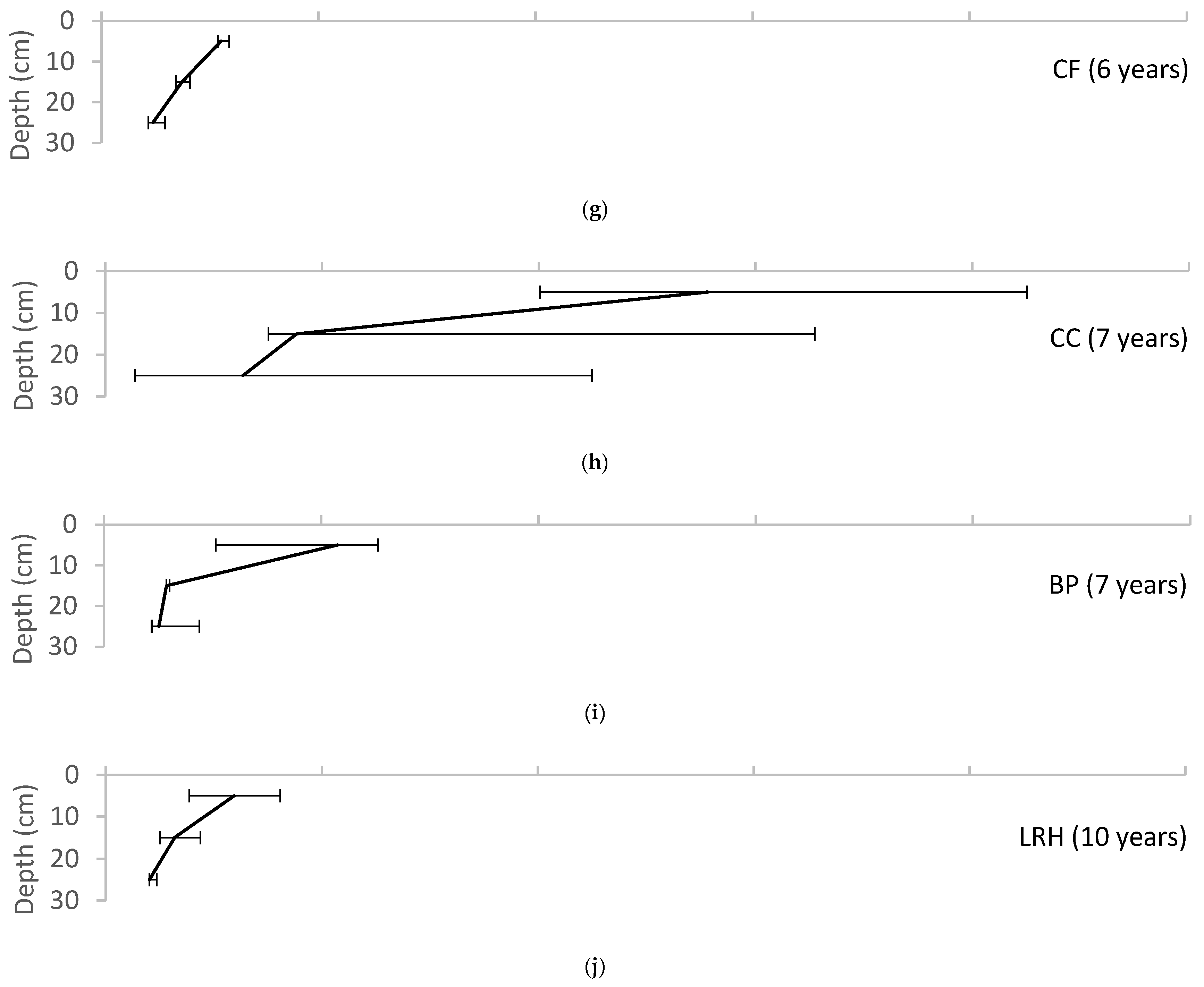
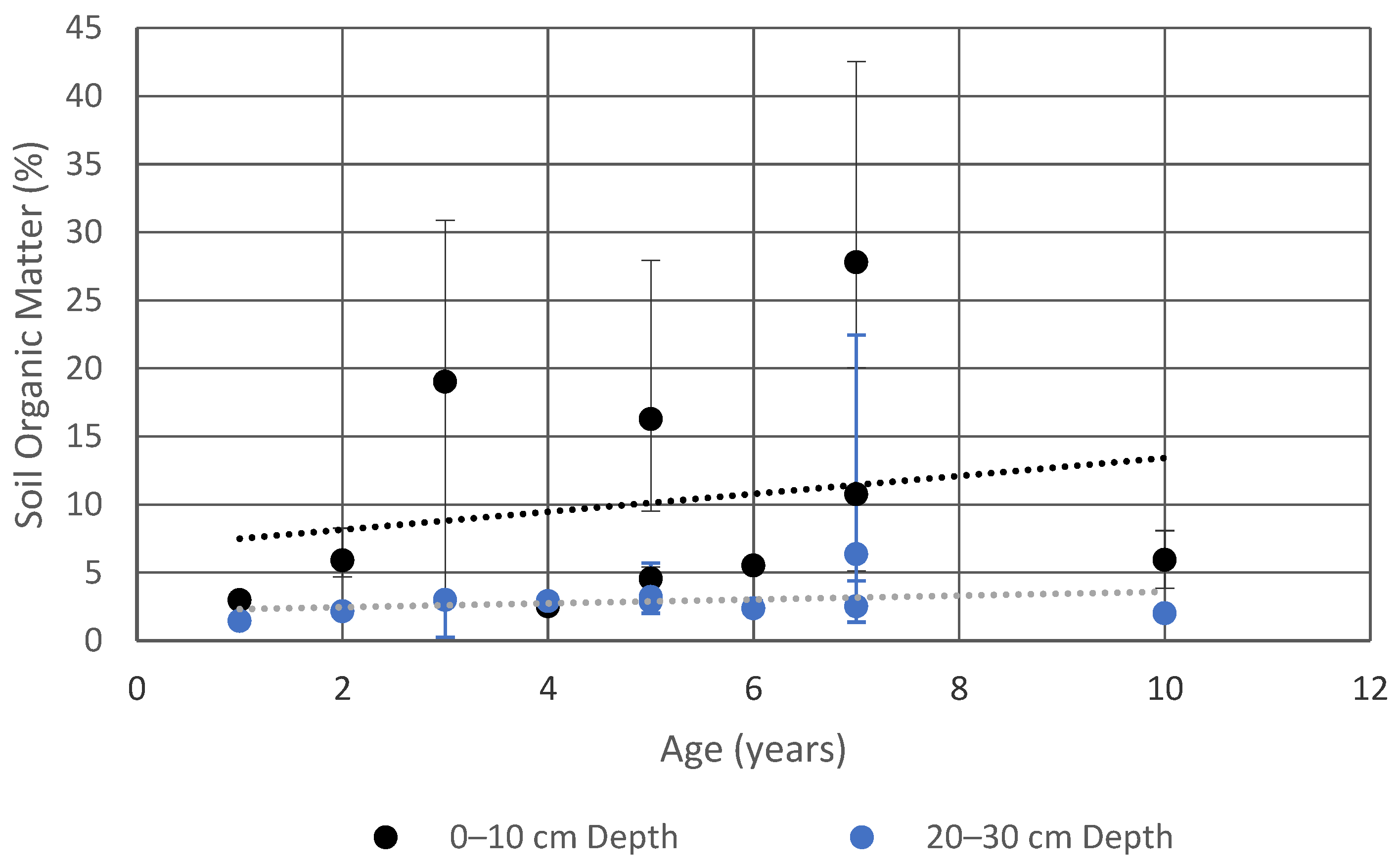
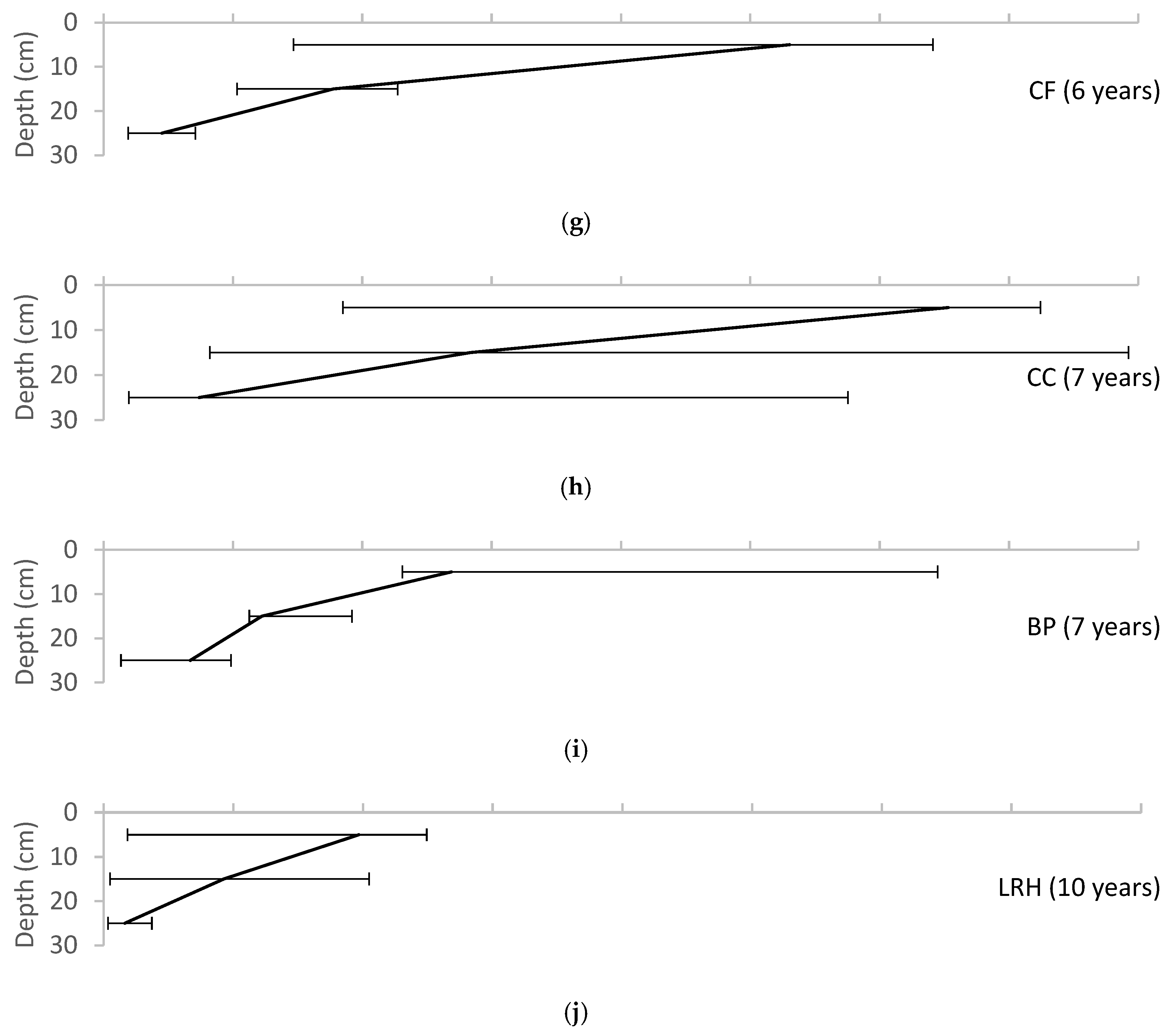
Soil organic matter was highest in the surface layer of the pits and declined with depth at all sites except for the Chesapeake Bay Foundation site (CBF), where it was low throughout the profile but showed a slight increase at the 20–30 cm depth (Figure 3). Soil organic matter at the seven-year-old site at the Chevy Chase Bank (CC) was very high at all depths, with 28% in the surface layer (0–10 cm), 9% in the subsurface layer (10–20 cm) and 6% in the base layer (20–30 cm). This site was unusual with the 6% content in the base layer being greater than the content in the base layer at most of the other sites, which ranged from 1–3%. The initial soil specification at this site is unknown, but during sampling the soil was observed to have much more mulch incorporated into the soil at all depths than was observed at the other sites. Figure 4 shows a trend of increasing soil organic matter with age in the uppermost soil layer, compared with fairly consistent organic content in the base layer.
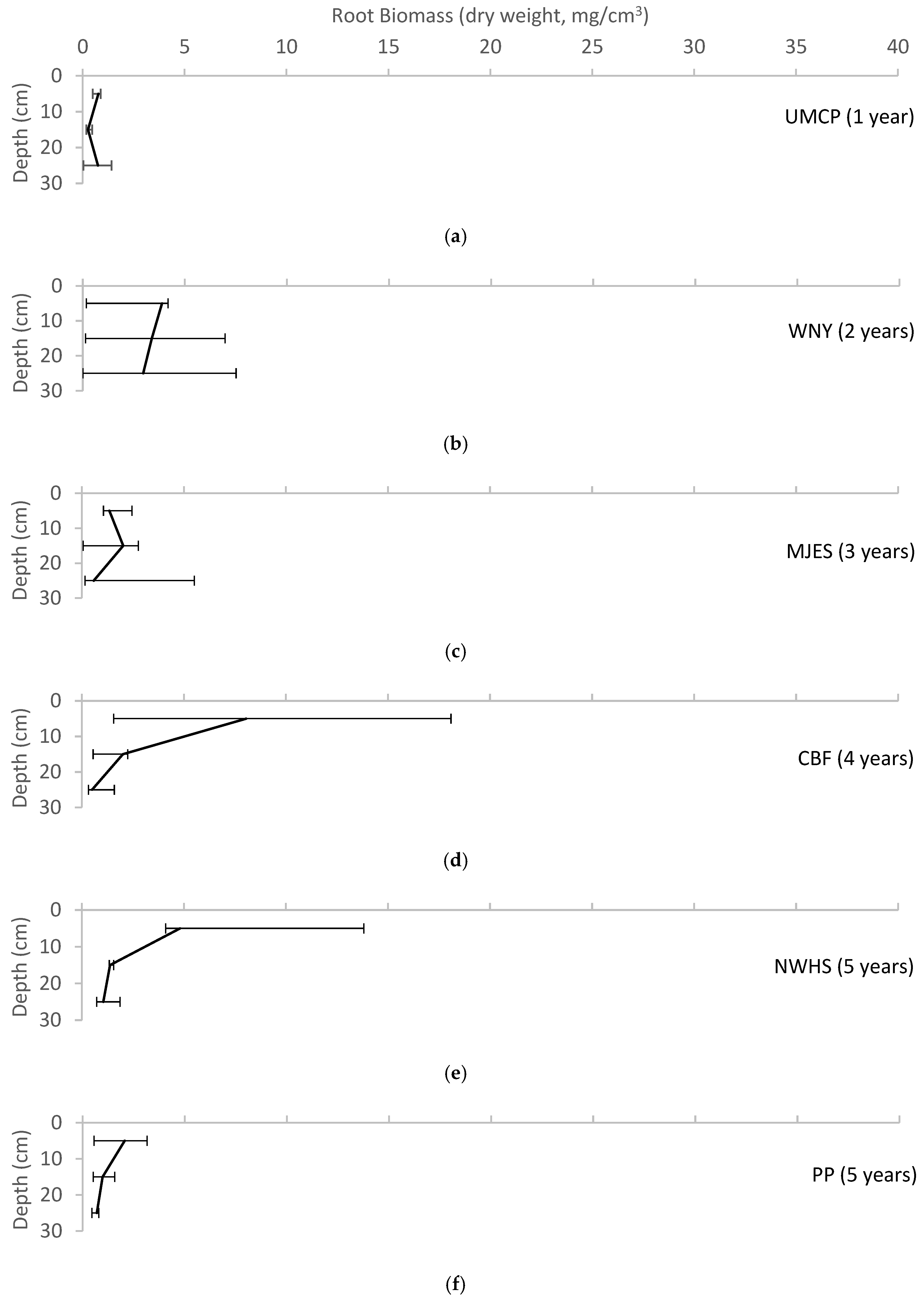
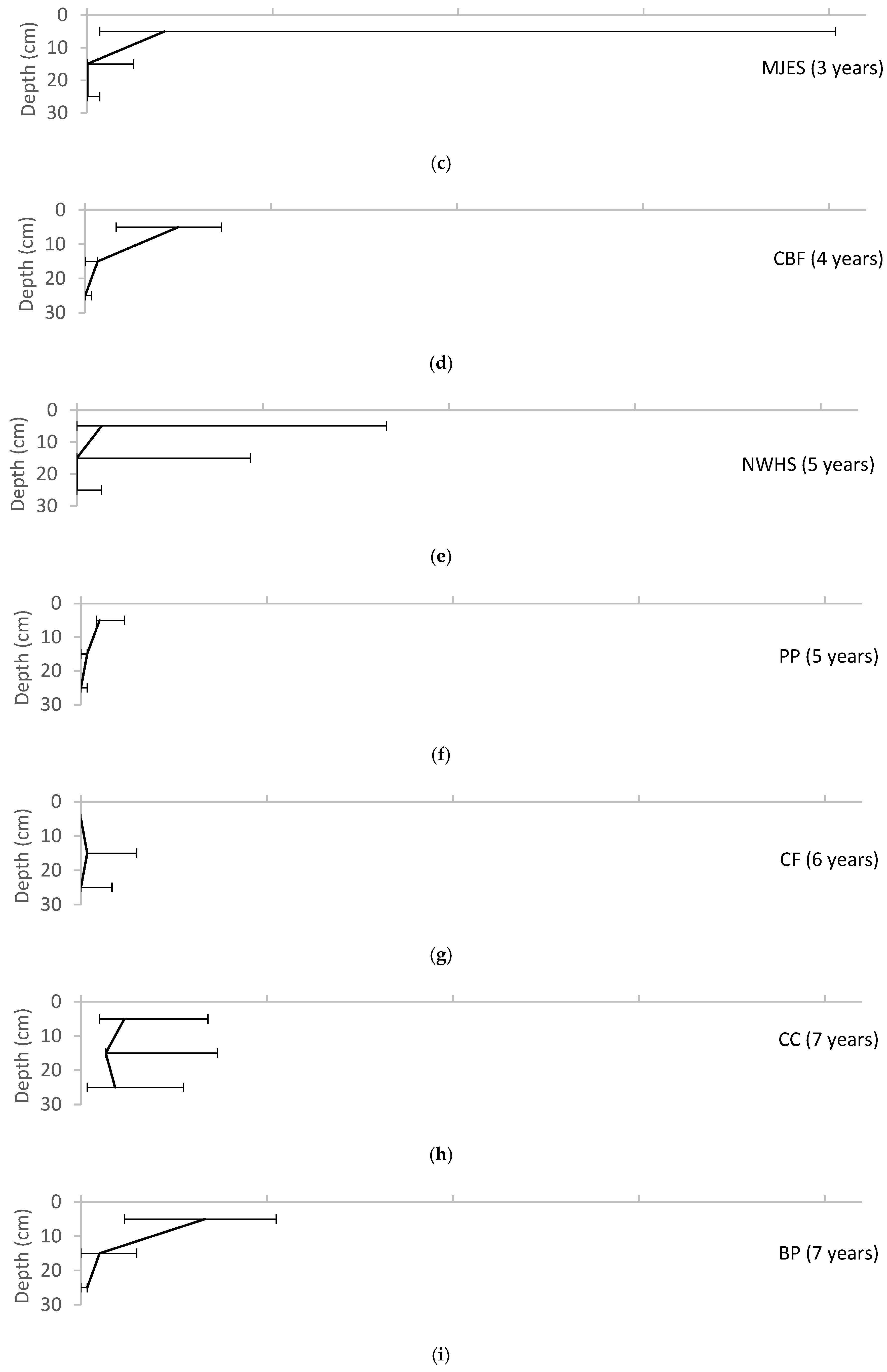
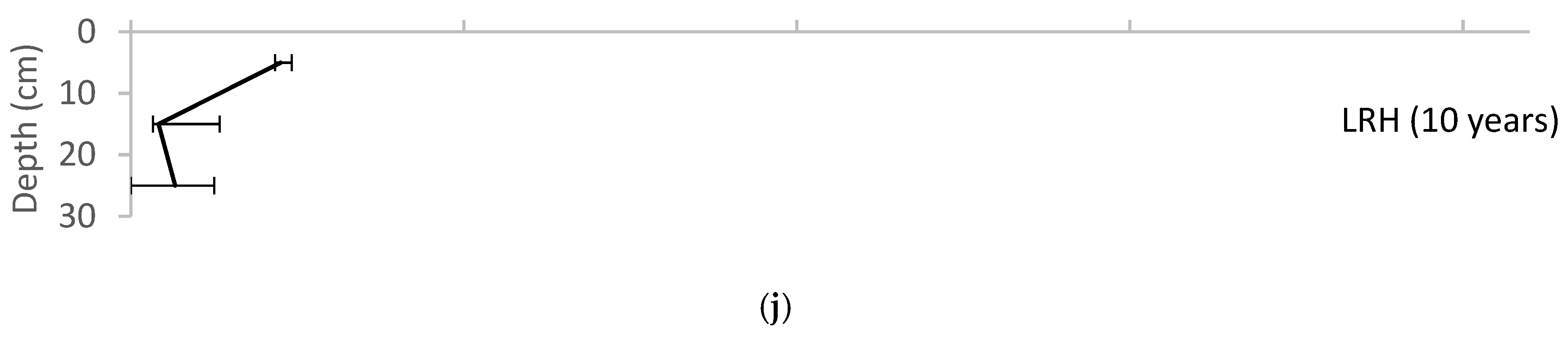
The root data showed the clearest view of increasing values with site age (Figure 5). The older sites (CF, CC, BP, and LRH) had relatively high average root biomass and a greater range of biomass from the surface to the base layers in the pits in comparison with root biomass at the youngest sites (UMCP, WNY, MJES).
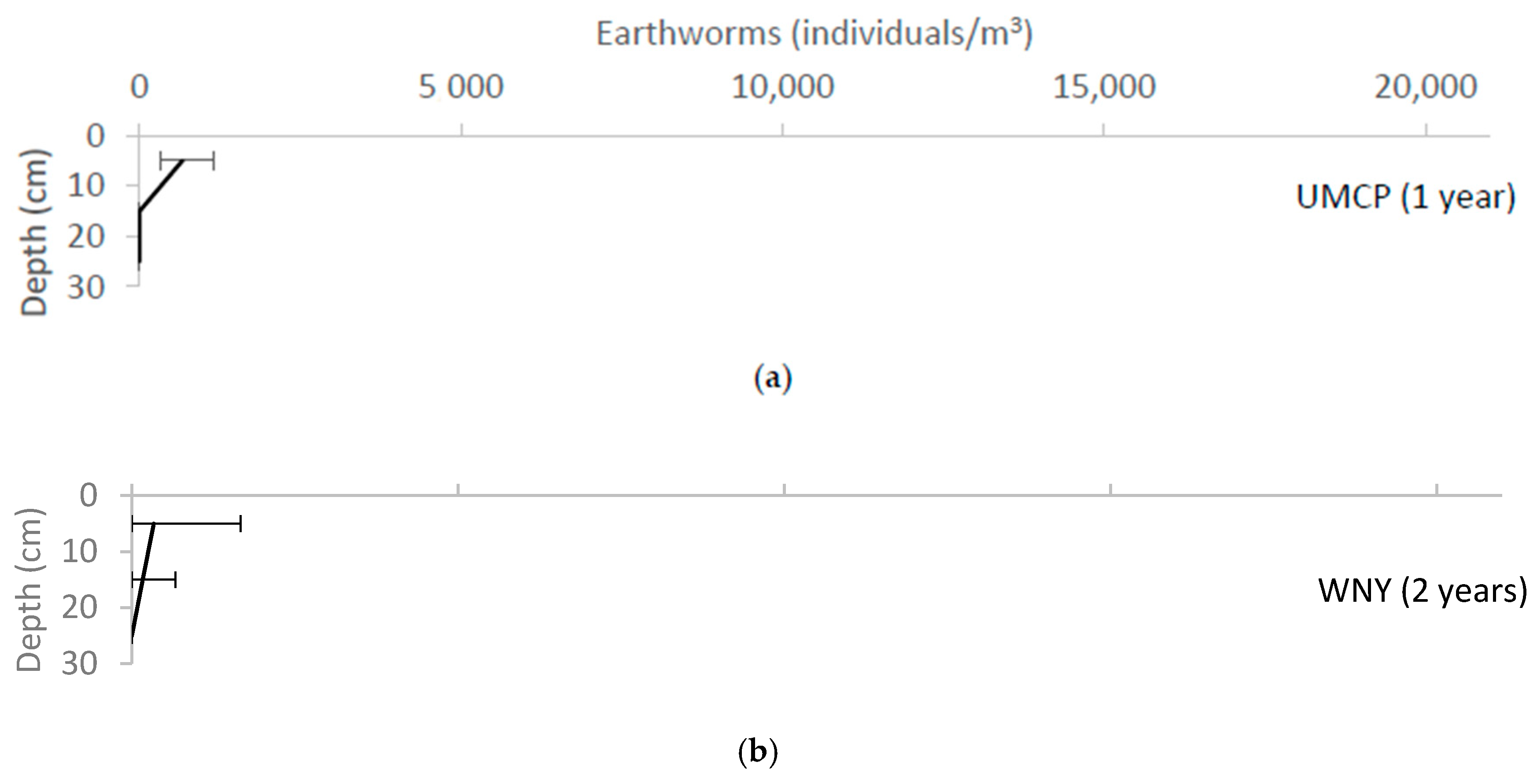
Earthworms were found at all 10 sites (Figure 6). Even the youngest site, UMCP, which had only been in operation for one year, and had the sandiest soil, had earthworms. In fact, earthworms are the only soil animal that was observed at all sites. Identified earthworm species included: Allolobophora chlorotica, Aporrectodea caliginosa, Bimastos parvus, Dendrodrilus ribidus, Diplocardia singularis, Eisenia fetida, Lumbricus rubellus, Lumbricus sp., Pheretima diffringens, and Pheretima sp.
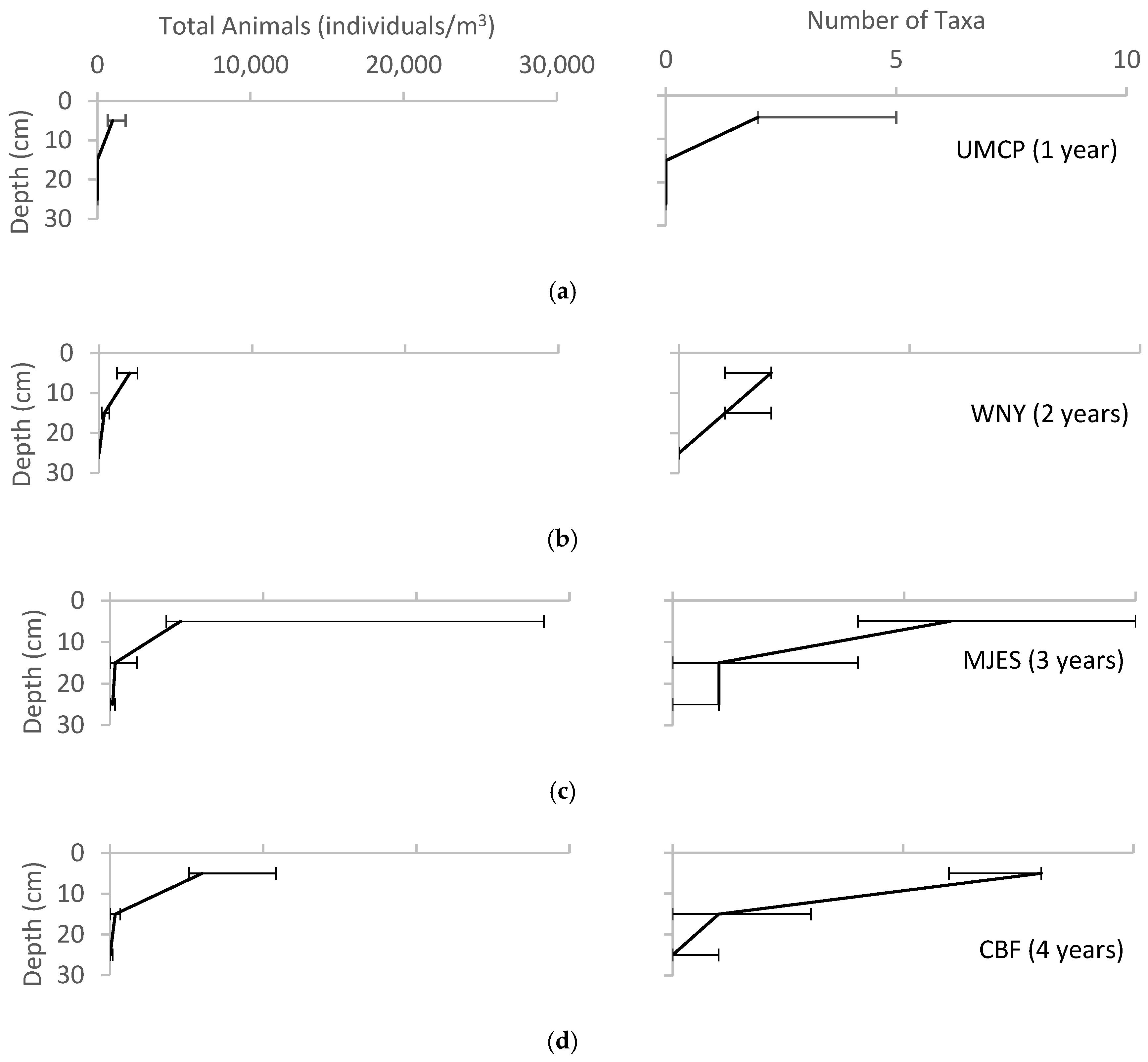
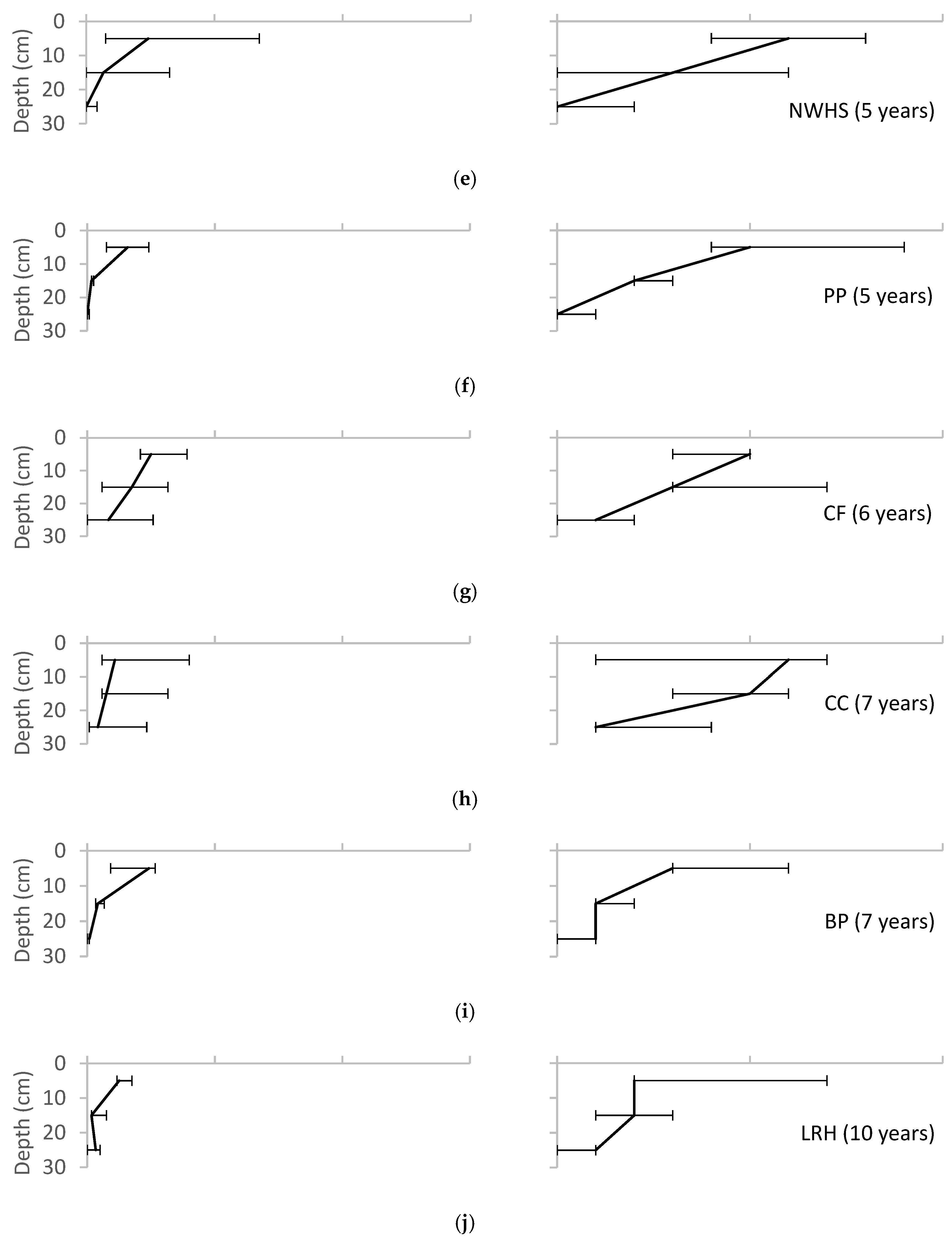
The absolute values of macroinvertebrate density and diversity varied greatly across sites, with the highest values found in the surface layers and declining values with depth (Figure 7). Earthworms (Oligochaeta, family Lumbricidae) were the most abundant macroinvertebrate group represented along with ants (Hymenoptera, family Formicidae) and adult beetles (Coleoptera). Other groups found in the pits included: beetle larvae (Coleoptera, family Scarabaeidae), centipedes (Chilopoda), millipedes (Diplopoda), pill bugs (Isopoda), pot worms (Enchytraeidae), spiders (Araneae), springtails (Collembola), slugs and snails (Gastropoda), fly larvae (Diptera), and mites (Acari), as shown in Table 3. Detailed data for all soil parameters at the sites are given in Ayers [38].
4. Discussion
The field surveys were intended to assess the extent of biological activity in the soil, to collect baseline soil physical data, and to look for evidence of pedogenesis in existing bioretention. The dataset shows a trend of increasing fines over time, as well as increased biological activity in the uppermost soil layer, indicated by elevated soil organic matter, plant roots, earthworms, and soil animals.
4.1. Baseline Physical Data
4.1.1. Soil Texture
Initial media specifications were available for 7 of the 10 field sites (Table 1). Unfortunately, comparison to testing results is confounded by two factors: first, media mixes are specified on a volume basis, and soil testing results are reported on a mass basis; and second, comparison would ideally be between current measurements and testing data from the as-built media rather than the specification, and these testing data were not available for any of the sites.
Prior to 2000, a statewide bioretention specification had not yet been established in Maryland, and there may have been much more variety in what media were used in bioretention. In general, early bioretention specifications had less stringent limits on which United States Department of Agriculture (USDA) soil textural classifications were acceptable for topsoil. Beltway Plaza Mall’s 1997 plans specified five parts topsoil to one part wet loose peat moss or rotted manure. The topsoil could be composed of loam, sandy loam, clay loam, silt loam, sandy clay loam, or loamy sand, and therefore permitted use of a topsoil with a much higher clay content than would be allowed under a more recent specification. The current State of Maryland bioretention specifications, which were established in 2000, specify a soil mix that is 50% sand, 30% topsoil (sandy loam or loamy sand), and 20% organic material in the form of compost or shredded hardwood mulch [19]. Therefore, differences in the particle size distribution of the original soil mix could account for some of the differences in soil textures measured in this study.
Nevertheless, when plotted as a chronosequence, the field sites do exhibit a trend toward increasing fines over time (Figure 2). This is likely to be, at least in part, the effect of years of enrichment of the system with fine material carried in by stormwater.
4.1.2. Soil Organic Matter Enrichment
Soil organic matter was measured by the loss-on-ignition method [20], which measures the total organic matter fraction smaller than 2 mm in diameter. This method, therefore, does not include large, undecomposed organic matter fragments. Bioretention soil media frequently use shredded hardwood mulch as an organic matter source, and much of this organic matter would be excluded from the measurement. This is likely appropriate, as organic matter pieces this large do not play the same role as smaller, more decomposed organic matter fractions. More problematically, the method does not distinguish between small, undecomposed particles and the more active, humified organic matter fractions which are of interest due to their prominent role in the maintenance of soil structure and their strong affinity for cations [2,39]. Measurement of this active fraction falls to future researchers. Some sites did contain a large amount of hardwood mulch in the soil samples, which had often degraded sufficiently to create many very small mulch pieces. This tended to drive up the measured soil organic matter content. Indeed, the three sites with elevated organic content, MJES, CC, and PP, were observed to have relatively large quantities of mulch mixed into the upper soil layers.
4.2. Soil Biota
The extent of the root systems present in these bioretention cells is an important measure of the level of biological activity in the soil. Roots are the backbone of the soil ecosystem, playing vital structural and functional roles [40]. As they grow and die back, roots create void space in the soil. Root exudates glue soil particles together into aggregates. Root cells slough off and die back, providing a major source of organic matter for micro- and mesofauna [16]. Root biomass was expected to increase with the age of the bioretention, and this is shown in Figure 2. The trend toward increasing root biomass with age is not monotonic, but the data suggest that older bioretention tend to have greater biomass than younger bioretention. This analysis is of course somewhat complicated by variations in vegetation between sites.
This study has revealed the basic structure of the soil animal community. A robust community of fauna was found, with representatives of 12 different invertebrate taxa (Table 3). The youngest sites (UMCP and WNY) had a much lower diversity than more established sites, as is expected considering that soil animals must colonize the bioretention cell by dispersion from nearby populations. Each of the fauna found plays a role in the development of the soil. The first step of pedogenesis, development of an organically-enriched A horizon, is fundamentally a biological process [16,41]. Plant litter falls on the soil surface. Millipedes, pill bugs, ants, and earthworms fragment this litter, mix it with bacteria in their guts, and incorporate this inoculated plant material into the soil matrix. Earthworms, pot worms, springtails, and mites consume this pre-digested plant material, further fragmenting it, and further boosting bacterial and fungal activity. Bacteria and fungi complete the process of humification, converting this decayed plant material into stable humus. Predators, such as centipedes, spiders, and some ants, regulate invertebrate populations, preventing the explosive growth of a population, which would lead to collapse. Invertebrates act as earth-movers as well. Burrowing animals, such as earthworms, ants, and beetle larvae, mix soil layers, incorporate organic matter into the soil, aerate the soil, improve infiltration, and create and destroy aggregates. Earthworms, snails, and slugs exude mucilage which acts to glue soil particles together [40].
4.3. Evidence of Pedogenesis
When treated as a chronosequence, the 10 field sites yield a picture of the development of a bioretention over a decade. Standard practice in the construction of bioretention cells is to uniformly mix the soil media, and then top the media with mulch to a depth of several inches. This would correspond to the starting point of Figure 1, in which an undifferentiated soil (the C horizon), is overlain by an organic (O) horizon. In these first 10 years, we see the beginnings of pedogenesis, with the gradual formation of an A horizon enriched with organic matter at the interface between the O and C horizons. Figure 4 and Figure 5 show the gradual development of an enriched organic layer in the uppermost soil layer (0–10 cm) over time. The profiles show both a shift toward higher organic matter in the upper layer and a shift toward a steeper gradient of organic matter between the upper and lower soil layers. A very similar pattern was seen by Leisman [10] in a chronosequence on abandoned mine spoil banks and is characteristic of early pedogenesis. The elevated soil organic matter observed in the soil profiles is evidence that this surface mulch layer is being decomposed and incorporated into the soil surface by earthworms and other soil animals.
Root biomass increases as plants become established; therefore, root biomass is expected to increase over the first years after planting. Figure 5 suggests a shift to the right with the age of the bioretention. The diagram shows the gradual formation of a gradient with depth, with the greatest root biomass in the topsoil, and decreasing at deeper soil levels. This is an expected result, corresponding to declining oxygen levels at greater soil depths.
Figure 6 suggests an increase in earthworm abundance over time at all depths, with greater increases in the uppermost soil layer. This is consistent with the gradual colonization of the sites by earthworms washed in from surrounding natural areas. It is to be noted that the MJES site had many more earthworms than any of the other sites. The reason for this is unknown.
Table 3 suggests increasing numbers of macroinvertebrates with bioretention age. All sites showed higher numbers of macroinvertebrates in the upper soil layers. Increasing species richness and density are expected patterns of the gradual colonization of a newly disturbed site. Earthworms had colonized all sites, including the youngest site, UMCP, which had only been in operation for one year. This suggests that earthworms may be an important pioneer species in the bioretention ecosystem.
4.4. Confounding Factors
Variations in the histories of these sites make them an imperfect chronosequence. They differ in many respects, some of which may influence organic matter, root biomass, and animal populations to a greater degree than the passage of time. The most important of these factors are the following: variations in soil texture, differences in plant cover, differences in the litter/mulch layer, and variations in the proximity to natural areas. It is well known that earthworms dislike very sandy soils [42]. This is thought to be due in part to the abrasive effects of sands on their soft bodies. The type and extent of plant cover can be expected to have a major influence on many aspects of the biology of the bioretention. Plant cover will determine to a large extent the root biomass that can be expected to develop over time. It will also determine the amount of plant litter that will accumulate on the bioretention surface, contributing to the organic content of the soil. The type of plants present can affect soil animals, as the litter produced may be more or less easily broken down. Proximity of bioretention to natural areas is likely to be an important factor in determining how quickly it will be colonized by soil invertebrates and may also determine which animals are likely to be introduced. Various management schedules could have an influence on biological development. For example, mowing, sediment removal and mulch replacement, use of pesticides and herbicides within the watershed, and use of road salts within the watershed would be expected to impact both plant and animal populations. Unfortunately, detailed data on the management of these sites are unavailable. Stevens and Walker [43], however, contend that all chronosequence studies fail in one way or another to control for all soil forming factors apart from time, but they assert that even flawed chronosequences can be useful for making qualitative comparisons.
4.5. Sources of Error in Animal Sampling
Sampling of soil-dwelling macroinvertebrates is notoriously difficult to perform accurately [44]. A number of factors could have influenced the survey results. Environmental factors, such as time of year, temperature on the sampling date, and the antecedent weather conditions, may have influenced macroinvertebrate populations at the time of sampling. In addition, the sampling method used, while appropriate for assessing the earthworm population, does not give an accurate population estimate for all taxa. Many of the very small soil animals, such as springtails, were probably missed during hand sorting. Ants tend to be heterogeneously spaced in colonies, interception of which may explain the anomalously high number of ants counted at CF (Table 3). For this reason, animal numbers should be used only for comparison between these sites and not for comparison with other studies.
4.6. Application to Bioretention Design
The organic component of bioretention media has the potential to both improve pollutant removal and act as a source of nutrient export [45]. This study does not address water quality directly, but the observed accumulation of organic matter in the top 10 cm of the bioretention soil may well have implications for pollutant removal performance. It may even make sense to simulate a naturally-occurring A horizon in bioretention design, confining the organic amendment to the top soil layer, the region where it is most available to plants and soil animals. This would allow for the lower soil layers to be customized to minimize nutrient leaching.
5. Conclusions
In order to develop new recommendations for bioretention maintenance, it is first necessary to understand how bioretention soils will behave over time. The current engineering design of bioretention assumes that the soil mix installed will remain largely unchanged for the lifetime of the cell. The only development anticipated is the accumulation of fines at the soil surface, which may need to be removed periodically. The principles of pedogenesis as understood by soil science suggest that these assumptions are inadequate. Once installed, bioretention will evolve in a manner similar to a disturbed soil in any other setting. That is, plants and soil animals will colonize the soil and will systematically change the soil structure. This survey has confirmed that soil-dwelling macroinvertebrates are ubiquitous in bioretention cells, in spite of their physical isolation. In particular, earthworms were found at all sites, and had colonized the sites within a year of construction. The field surveys show a clear development of a characteristic soil profile with decreasing soil organic matter and biological activity with depth.
Future research could help to identify what, if any, effect this biological activity has on hydraulic conductivity and/or pollutant removal.
Engineering a living system is a challenging task, requiring an understanding of both the physical requirements to be met and the ways in which the system will behave and evolve over time. This study is an attempt to integrate ecology and soil science with civil engineering. Bioretention is a novel kind of ecosystem and is not yet well characterized. Improving our understanding of bioretention ecology and how bioretention ecosystems interact with the physical properties of bioretention will allow us to design better systems.
Author Contributions
Conceptualization, E.M.A.; Formal analysis, E.M.A.; Investigation, E.M.A. and P.K.; Supervision, P.K.; Writing—original draft, E.M.A.; Writing—review and editing, P.K.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Johnson, D.; Watson-Stegner, D. Evolution model of pedogenesis. Soil Sci. 1987, 143, 349–366. [Google Scholar] [CrossRef]
- Brady, N.; Weil, R. The Nature and Properties of Soils, 13th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2002. [Google Scholar]
- Li, H.; Davis, A.P. Heavy metal capture and accumulation in bioretention media. Environ. Sci. Technol. 2008, 42, 5247–5253. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Davis, A.P. Urban particle capture in bioretention media. I: Laboratory and field studies. J. Environ. Eng. 2008, 134, 409–418. [Google Scholar] [CrossRef]
- Emerson, C.H.; Traver, R.G. Multiyear and seasonal variation of infiltration for storm-water best management practices. J. Irrig. Drain. Eng. 2008, 134, 598–605. [Google Scholar] [CrossRef]
- Gilbert Jenkins, J.K.; Wadzuk, B.M.; Welker, A.L. Fines accumulation and distribution in a storm-water rain garden nine years postconstruction. J. Irrig. Drain. Eng. 2010, 136, 862–869. [Google Scholar] [CrossRef]
- Le Coustumer, S.; Fletcher, T.D.; Deletic, A.; Barraud, S.; Lewis, J.F. Hydraulic performance of biofilter systems for stormwater management: Influences of design and operation. J. Hydrol. 2009, 376, 16–23. [Google Scholar] [CrossRef]
- Jenny, H. Factors of Soil Formation; McGraw-Hill: New York, NY, USA, 1941. [Google Scholar]
- Pickett, S.T. Space-for-time substitution as an alternative to long-term studies. In Long-Term Studies in Ecology: Approaches and Alternatives; Springer-Verlag: New York, NY, USA, 1989; pp. 110–135. [Google Scholar]
- Leisman, G.A. A vegetation and soil chronosequence on the Mesabi Iron Range Spoil Banks, Minnesota. Ecol. Monogr. 1957, 27, 221–245. [Google Scholar] [CrossRef]
- Kangas, P.C. Succession as an alternative for reclaiming phosphate spoil mounds. In Studies on Phosphate Mining, Reclamation and Energy; Center for Wetlands, University of Florida: Gainesville, FL, USA, 1983. [Google Scholar]
- Frouz, J.; Keplin, B.; Pizl, V.; Tajovsky, K.; Stary, J.; Lukesov, A.; Novkov, A.; Balk, V.; Hnel, L.; Materna, J.; et al. Soil biota and upper soil layers development in two contrasting post-mining chronosequences. Ecol. Eng. 2001, 17, 275–284. [Google Scholar] [CrossRef]
- Gonzalez-Sangregorio, M.; Trasar-Cepeda, M.; Leiros, M.; Gil-Sotres, F.; Guitian-Ojea, F. Early stages of lignite mine soil genesis: Changes in biochemical properties. Soil Biol. Biochem. 1991, 23, 589–595. [Google Scholar] [CrossRef]
- Roberts, J.; Daniels, W.; Bell, J.; Burger, J. Early stages of mine soil genesis as affected by topsoiling and organic amendments. Soil Sci. Soc. Am. J. 1988, 52, 730–738. [Google Scholar] [CrossRef]
- Roberts, J.; Daniels, W.; Bell, J.; Burger, J. Early stages of mine soil genesis in a southwest Virginia spoil lithosequence. Soil Sci. Soc. Am. J. 1988, 52, 716–723. [Google Scholar] [CrossRef]
- Gobat, J.; Aragno, M.; Matthey, W. The Living Soil: Fundamentals of Soil Science and Soil Biology; Science Publishers: Enfield, NH, USA, 2003. [Google Scholar]
- Oades, J. The role of biology in the formation, stabilization and degradation of soil structure. Geoderma 1993, 56, 377–400. [Google Scholar] [CrossRef]
- Hoogerkamp, M.; Rogaar, H.; Eijsackers, H. Effect of earthworms on grassland on recently reclaimed polder soils in the Netherlands. In Earthworm Ecology: From Darwin to Vermiculture; Satchell, J., Ed.; Chapman and Hall, Ltd.: London, UK, 1983; pp. 85–105. [Google Scholar]
- Maryland Department of the Environment. Maryland Stormwater Design Manual; State of Maryland: Baltimore, MD, USA, 2000.
- American Society for Testing and Materials. Standard Test Methods for Moisture, Ash, and Organic Matter of Peat and Other Organic Soils; Technical Report ASTM D2974-00; American Society for Testing and Materials: West Conshohocken, PA, USA, 2000. [Google Scholar]
- Csuzdi, C.; Slavecz, K. Lumbricus friendi Cognetti, 1904, a new exotic earthworm in North America. Northeast. Nat. 2003, 10, 77–82. [Google Scholar] [CrossRef]
- Reynolds, J. The earthworms of Maryland (Oligochaeta: Acanthodrilidae, Lumbricidae, Megasolecidae and Sparganophilidae). Megadrilogica 1974, 1, 1–12. [Google Scholar]
- Baker, G.; Barrett, V. Earthworm Identifier; CSIRO: Melbourne, Australia, 1994. [Google Scholar]
- Eaton, T. Earthworms of the northeastern United States: A key, with distribution records. J. Wash. Acad. Sci. 1942, 32, 242–249. [Google Scholar]
- Fender, W.M.; McKey-Fender, D. Oligochaeta: Megascolecidae and other earthworms from western North America. In Soil Biology Guide; Dindal, D.L., Ed.; John Wiley & Sons: New York, NY, USA, 1990; pp. 357–377. [Google Scholar]
- Gates, G. The genus Pheretima in North America. Bull. Mus. Comp. Zool. Harvard Univ. 1937, 80, 339–373. [Google Scholar]
- Gates, G. On some species of the oriental earthworm genus Pheretima Kinberg, 1867, with key to species reported from the Americas. Am. Mus. Novit. 1958, 1888, 1–33. [Google Scholar]
- Gates, G. Contributions to North American Earthworms (Annelida), No. 4: On American earthworm genera. I. Eisenoides (Lumbricidae). Bull. Tall Timbers Res. Stn. 1972, 3, 1–17. [Google Scholar]
- Gates, G. Contributions to North American Earthworms (Annelida), No. 5: On variation in another anthropochorous species of the oriental earthworm genus Pheretima Kinberg 1866 (Megascolecidae). Bull. Tall Timbers Res. Stn. 1972, 3, 18–44. [Google Scholar]
- Gates, G. Contributions to North American Earthworms (Annelida), No. 8: The Earthworm Genus Octolasion in America. Bull. Tall Timbers Res. Stn. 1973, 4, 29–50. [Google Scholar]
- Gates, G. Contributions to North American Earthworms (Annelida), No. 10: Contributions to a revision of the Lumbricidae. X., Dendrobaena octaedra (Savigny) 1826, with special reference to the importance of its parthenogenetic polymorphism for the classification of earthworms. Bull. Tall Timbers Res. Stn. 1974, 5, 15–57. [Google Scholar]
- James, W.M. Oligochaeta: Megascolecidae and other earthworms from southern and midwestern North America. In Soil Biology Guide; Dindal, D.L., Ed.; John Wiley & Sons: New York, NY, USA, 1990; pp. 379–386. [Google Scholar]
- Lee, K. The Earthworm Fauna of New Zealand; New Zealand Department of Scientific and Industrial Research: Wellington, New Zealand, 1959.
- Reynolds, J.W.; Clebsch, E.E.C.; Reynolds, W.M. Contributions to North American Earthworms (Oligochaeta), No. 13: The Earthworms of Tennessee (Oligochaeta). I. Lumbricidae. Bull. Tall Timbers Res. Stn. 1974, 3, 1–133. [Google Scholar]
- Schwert, D.P. Oligochaeta: Lumbricidae. In Soil Biology Guide; Dindal, D.L., Ed.; John Wiley & Sons: New York, NY, USA, 1990; pp. 341–356. [Google Scholar]
- Sims, R.W.; Gerard, B.M.; Barnes, R.S.K.; Crothers, J.H. Earthworms. Synopses of the British Fauna; Linnean Society of London: Shrewsbury, UK, 1999. [Google Scholar]
- Worm Watch Canada Field Guide to Earthworms. Available online: https://www.naturewatch.ca/wormwatch/how-to-guide/identifying-earthworms/ (accessed on 5 November 2018).
- Ayers, E.M. Pedogenesis in Rain Gardens: The Role of Earthworms and Other Organisms in Long-Term Soil Development. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2009. [Google Scholar]
- Weil, R.R.; Islam, K.R.; Stine, M.A.; Gruver, J.B.; Samson-Liebig, S.E. Estimating active carbon for soil quality assessment: A simplified method for lab and field use. Am. J. Altern. Agric. 2003, 18, 3–17. [Google Scholar]
- Six, J.; Bossuyt, J.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Hole, F. Effects of animals on the soil. Geoderma 1981, 25, 75–112. [Google Scholar] [CrossRef]
- Curry, J. Factors affecting earthworm abundance in soils. In Earthworm Ecology; Edwards, C., Ed.; St. Lucie Press: Boca Raton, FL, USA, 1998; pp. 37–64. [Google Scholar]
- Stevens, P.; Walker, T. The chronosequence concept and soil formation. Q. Rev. Biol. 1970, 45, 333–350. [Google Scholar] [CrossRef]
- Southwood, T.; Henderson, P. Ecological Methods, 3rd ed.; Blackwell Science Ltd.: Malden, MA, USA, 2000. [Google Scholar]
- LeFevre, G.H.; Paus, K.H.; Natarajan, P.; Gulliver, J.S.; Novak, P.J.; Hozalski, R.M. Review of dissolved pollutants in urban storm water and their removal and fate in bioretention cells. J. Environ. Eng. 2014, 141, 04014050. [Google Scholar] [CrossRef]
Figure 1.
Conceptual illustration of the initial stages of pedogenesis, after Brady and Weil [2]. An organic (O) horizon forms at the soil surface (commonly referred to as the “litter layer”) and is gradually broken down and incorporated into the uppermost soil layer, forming an A horizon (commonly referred to as “topsoil”).
Figure 1.
Conceptual illustration of the initial stages of pedogenesis, after Brady and Weil [2]. An organic (O) horizon forms at the soil surface (commonly referred to as the “litter layer”) and is gradually broken down and incorporated into the uppermost soil layer, forming an A horizon (commonly referred to as “topsoil”).

Figure 2.
Fines content of bioretention media vs. time. Fines are defined as the sum of the silt and clay fractions. The value for each field site represents the mean value for all samples at all depths. Error bars represent the standard deviation from the mean.
Figure 2.
Fines content of bioretention media vs. time. Fines are defined as the sum of the silt and clay fractions. The value for each field site represents the mean value for all samples at all depths. Error bars represent the standard deviation from the mean.

Figure 3.
Variation of soil organic matter with depth for each of the field sites, reported as median ± range. (a) UMCP; (b) WNY; (c) MJES; (d) CBF; (e) NWHS; (f) PP; (g) CF; (h) CC; (i) BP; (j) LRH.
Figure 3.
Variation of soil organic matter with depth for each of the field sites, reported as median ± range. (a) UMCP; (b) WNY; (c) MJES; (d) CBF; (e) NWHS; (f) PP; (g) CF; (h) CC; (i) BP; (j) LRH.


Figure 4.
Soil organic matter vs. time for the upper (0–10 cm) and lowest (20–30 cm) soil layers sampled, reported as median ± range.
Figure 4.
Soil organic matter vs. time for the upper (0–10 cm) and lowest (20–30 cm) soil layers sampled, reported as median ± range.

Figure 5.
Variation of root biomass with depth for each of the field sites, reported as median ± range. (a) UMCP; (b)WNY; (c) MJES; (d) CBF; (e) NWHS; (f) PP; (g) CF; (h) CC; (i) BP; (j) LRH.
Figure 5.
Variation of root biomass with depth for each of the field sites, reported as median ± range. (a) UMCP; (b)WNY; (c) MJES; (d) CBF; (e) NWHS; (f) PP; (g) CF; (h) CC; (i) BP; (j) LRH.


Figure 6.
Variation of earthworm density with depth for each of the field sites, reported as median ± range. (a) UMCP; (b)WNY; (c) MJES; (d) CBF; (e) NWHS; (f) PP; (g) CF; (h) CC; (i) BP; (j) LRH.
Figure 6.
Variation of earthworm density with depth for each of the field sites, reported as median ± range. (a) UMCP; (b)WNY; (c) MJES; (d) CBF; (e) NWHS; (f) PP; (g) CF; (h) CC; (i) BP; (j) LRH.



Figure 7.
Variation of total animals and taxa found with depth for each of the field sites, reported as median ± range. (a) UMCP; (b)WNY; (c) MJES; (d) CBF; (e) NWHS; (f) PP; (g) CF; (h) CC; (i) BP; (j) LRH.
Figure 7.
Variation of total animals and taxa found with depth for each of the field sites, reported as median ± range. (a) UMCP; (b)WNY; (c) MJES; (d) CBF; (e) NWHS; (f) PP; (g) CF; (h) CC; (i) BP; (j) LRH.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Field sites. Bioretention media specification represents the specification as included in construction documents, where available.
Table 1.
Field sites. Bioretention media specification represents the specification as included in construction documents, where available.
| Site Identifier | Location | Year of Construction | Age at Sampling (Years) | Bioretention Media Specification (Volume Basis) |
|---|---|---|---|---|
| UMCP | University of Maryland College Park, MD | 2004 | 1 | 50% sand |
| 30% topsoil—sandy loam or loamy sand | ||||
| 20% shredded 2 × hardwood mulch | ||||
| WNY | Washington Navy Yard Washington, DC | 2002 | 2 | 50% sand |
| 30% topsoil—sandy loam or loamy sand | ||||
| 20% shredded 2 × hardwood mulch | ||||
| MJES | Mary Harris ”Mother” Jones Elementary School Adelphi, MD | 2002 | 3 | 50% sand |
| 25% topsoil—sandy loam or loam | ||||
| 25% compost | ||||
| CBF | Philip Merrill Environmental Center Annapolis, MD | 2001 | 4 | 60–65% loamy sand |
| 35–40% compost | ||||
| OR | ||||
| 30% sandy loam | ||||
| 30% coarse sand | ||||
| 40% compost 1 | ||||
| NWHS | Northwestern High School Hyattsville, MD | 1999 | 5 | 50% sand |
| 25% topsoil | ||||
| 25% compost | ||||
| PP | Inglewood Center III Upper Marlboro, MD | 1999 | 5 | 70% sand |
| 30% compost | ||||
| CF | Claggett Farm Upper Marlboro, MD | 1999 | 6 | Unavailable |
| CC | Chevy Chase Bank Silver Spring, MD | 1998 | 7 | Unavailable |
| BP | Beltway Plaza Mall Greenbelt, MD | 1997 | 7 | 83% topsoil—loam, sandy loam, clay loam, silt loam, sandy clay loam, or loamy sand |
| 17% peat moss or rotted manure | ||||
| LRH | Laurel Regional Hospital Laurel, MD | 1994 | 10 | Unavailable |
1 Construction documents unavailable. Media specification is assumed to conform to the Maryland Department of the Environment’s standard bioretention media specification [19], (p. B.4.7), established in 2000.
Table 2.
Particle size analysis, separated into United States Department of Agriculture (USDA) textural classes. Reported as median (range).
Table 2.
Particle size analysis, separated into United States Department of Agriculture (USDA) textural classes. Reported as median (range).
| Site | Sand (%) | Silt (%) | Clay (%) | d60/d10 |
|---|---|---|---|---|
| Surface (0–10 cm) | ||||
| UMCP | 87.7 (88.3–92.1) | 7.3 (2.8–8.2) | 4.3 (4.0–5.1) | 10 |
| WNY | 71.8 (68.1–75.0) | 21.7 (21.0–26.4) | 5.4 (3.9–6.5) | 10 |
| MJES | 49.9 (43.1–78.7) | 31.8 (13.2–41.3) | 15.6 (8.1–18.3) | 50 |
| CBF | 60.6 (59.8–62.6) | 14.5 (14.0–16.9) | 22.9 (22.5–26.2) | 50 |
| NWHS | 51.3 (50.3–57.6) | 32.1 (32.1–34.7) | 15.0 (10.3–16.6) | 50 |
| PP | 69.6 (62.9–79.1) | 13.2 (8.3–17.5) | 17.2 (12.6–19.6) | 50 |
| CF | 69.9 (67.5–72.6) | 17.9 (16.6–18.0) | 12.1 (10.8–14.5) | 50 |
| CC | 67.0 (43.8–76.4) | 28.3 (21.3–43.5) | 4.6 (2.3–12.7) | 5 |
| BP | 60.0 (53.7–72.8) | 17.9 (10.6–20.1) | 22.1 (16.6–26.2) | 50 |
| LRH | 59.2 (22.6–59.4) | 21.2 (20.7–46.0) | 17.5 (16.4–20.1) | 50 |
| Sub-Surface (10–20 cm) | ||||
| UMCP | 91.4 (91.4–93.8) | 4.0 (4.0–5.1) | 2.2 (2.2–3.6) | 10 |
| WNY | 72.4 (67.9–75.6) | 19.7 (17.1–26.8) | 7.3 (5.2–7.9) | 10 |
| MJES | 60.4 (57.2–96.3) | 21.0 (1.2–21.2) | 18.6 (2.5–21.6) | 50 |
| CBF | 60.8 (59.8–61.8) | 17.4 (16.5–17.5) | 22.7 (20.7–22.8) | 50 |
| NWHS | 40.1 (38.1–51.9) | 42.4 (31.0–46.7) | 17.1 (13.2–19.5) | 20 |
| PP | 87.9 (83.8–88.1) | 4.3 (4.2–11.9) | 7.6 (4.3–7.9) | 10 |
| CF | 67.5 (65.1–70.2) | 18.5 (16.8–19.8) | 13.0 (12.7–16.4) | 50 |
| CC | 48.0 (32.3–81.4) | 37.0 (15.3–56.1) | 11.6 (3.3–15.0) | 50 |
| BP | 54.9 (53.3–55.1) | 23.4 (21.1–24.0) | 22.7 (21.5–24.0) | 50 |
| LRH | 39.0 (35.6–62.4) | 25.4 (22.0–46.1) | 18.2 (12.1–39.0) | 20 |
| Sub-Sub-Surface (20–30 cm) | ||||
| UMCP | 92.0 (91.3–92.6) | 4.1 (3.8–5.4) | 3.6 (3.3–3.9) | 10 |
| WNY | 68.9 (66.5–75.2) | 25.6 (7.0–26.8) | 6.7 (5.5–17.8) | 10 |
| MJES | 57.2 (53.1–96.2) | 19.9 (1.6–25.8) | 21.1 (2.2–22.9) | 50 |
| CBF | 61.4 (60.6–61.8) | 16.9 (16.5–18.3) | 21.7 (19.9–22.9) | 50 |
| NWHS | 52.7 (52.1–68.5) | 30.9 (24.0–33.9) | 14.0 (7.5–16.4) | 50 |
| PP | 89.2 (87.8–92.2) | 4.2 (1.5–7.5) | 6.3 (3.3–8.0) | 10 |
| CF | 62.8 (61.2–64.9) | 17.4 (16.8–18.0) | 19.2 (17.7–22.0) | 20 |
| CC | 53.1 (46.7–93.8) | 31.0 (3.8–39.1) | 14.2 (2.4–15.9) | 50 |
| BP | 44.8 (27.7–44.8) | 34.9 (21.1–38.6) | 23.8 (20.4–33.7) | 20 |
| LRH | 40.3 (22.6–59.4) | 40.1 (21.1–57.1) | 19.6 (19.5–20.3) | 20 |
Table 3.
Abundance of macroinvertebrates found at field sites. Values represent total number of individuals counted at site.
Table 3.
Abundance of macroinvertebrates found at field sites. Values represent total number of individuals counted at site.
| Taxonomic Group | UMCP | WNY | MJES | CBF | NWHS | PP | CF | CC | BP | LRH |
|---|---|---|---|---|---|---|---|---|---|---|
| Lumbricidae (Earthworms) | 13 | 17 | 145 | 47 | 86 | 16 | 16 | 84 | 52 | 65 |
| Coleoptera (Beetles—adults and larvae) | 5 | 24 | 24 | 24 | 19 | 38 | 25 | 12 | 3 | 5 |
| Formicidae (Ants) | 1 | - | 8 | 8 | 12 | 4 | 148 | 22 | 10 | 2 |
| Enchytraeidae (Pot Worms) | - | - | 26 | 26 | 34 | 3 | - | 3 | - | - |
| Collembola (Springtails) | - | - | 23 | 23 | 9 | 3 | 1 | 1 | - | - |
| Chilopoda (Centipedes) | - | - | 1 | 1 | 1 | - | 6 | 29 | 3 | 1 |
| Isopoda (Pill bugs) | - | - | - | - | 3 | 2 | 1 | 3 | 4 | 3 |
| Diplopoda (Millipedes) | 1 | - | 3 | 3 | 1 | - | 7 | - | 1 | 3 |
| Araneae (Spiders) | 1 | - | 2 | 2 | 5 | 1 | 4 | 2 | - | - |
| Acari (Mites) | - | - | - | - | - | - | - | 1 | - | - |
| Gastropoda (Snails) | - | - | 1 | - | - | - | - | - | - | - |
| Diptera (Flies—larvae) | - | - | - | - | - | - | - | - | - | 1 |
| Unidentified Insect Larvae | - | - | 2 | 2 | 1 | - | 2 | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mitchell Ayers, E.; Kangas, P. Soil Layer Development and Biota in Bioretention. Water 2018, 10, 1587. https://doi.org/10.3390/w10111587
AMA Style
Mitchell Ayers E, Kangas P. Soil Layer Development and Biota in Bioretention. Water. 2018; 10(11):1587. https://doi.org/10.3390/w10111587
Chicago/Turabian StyleMitchell Ayers, Emily, and Patrick Kangas. 2018. "Soil Layer Development and Biota in Bioretention" Water 10, no. 11: 1587. https://doi.org/10.3390/w10111587
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.