Quantifying Urban Bioswale Nitrogen Cycling in the Soil, Gas, and Plant Phases

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bioswale Sites
2.2. Soil
2.2.1. Soil Characterization
2.2.2. Soil Mineralization and Nitrification
2.3. Soil Gas Emissions
2.4. Plant Uptake
2.5. Statistical Analysis
3. Results
3.1. Soil
3.1.1. Soil Characterization
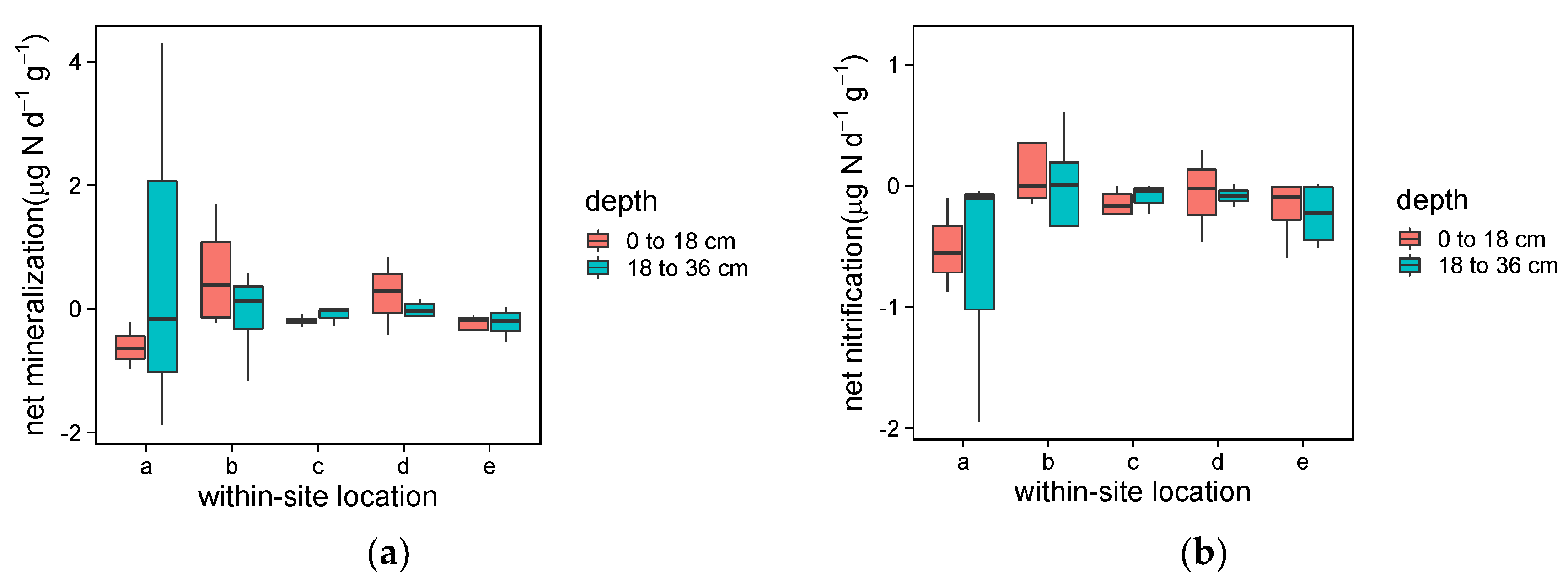
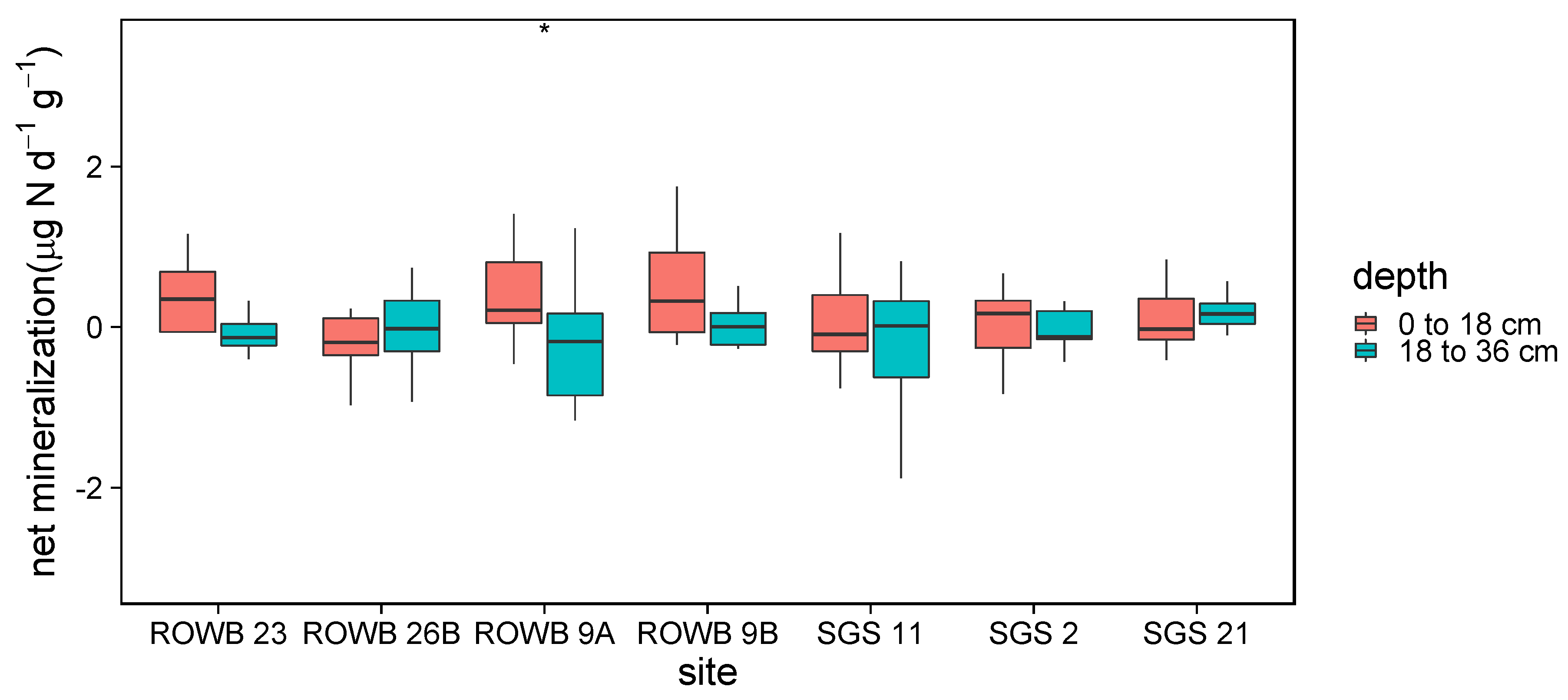
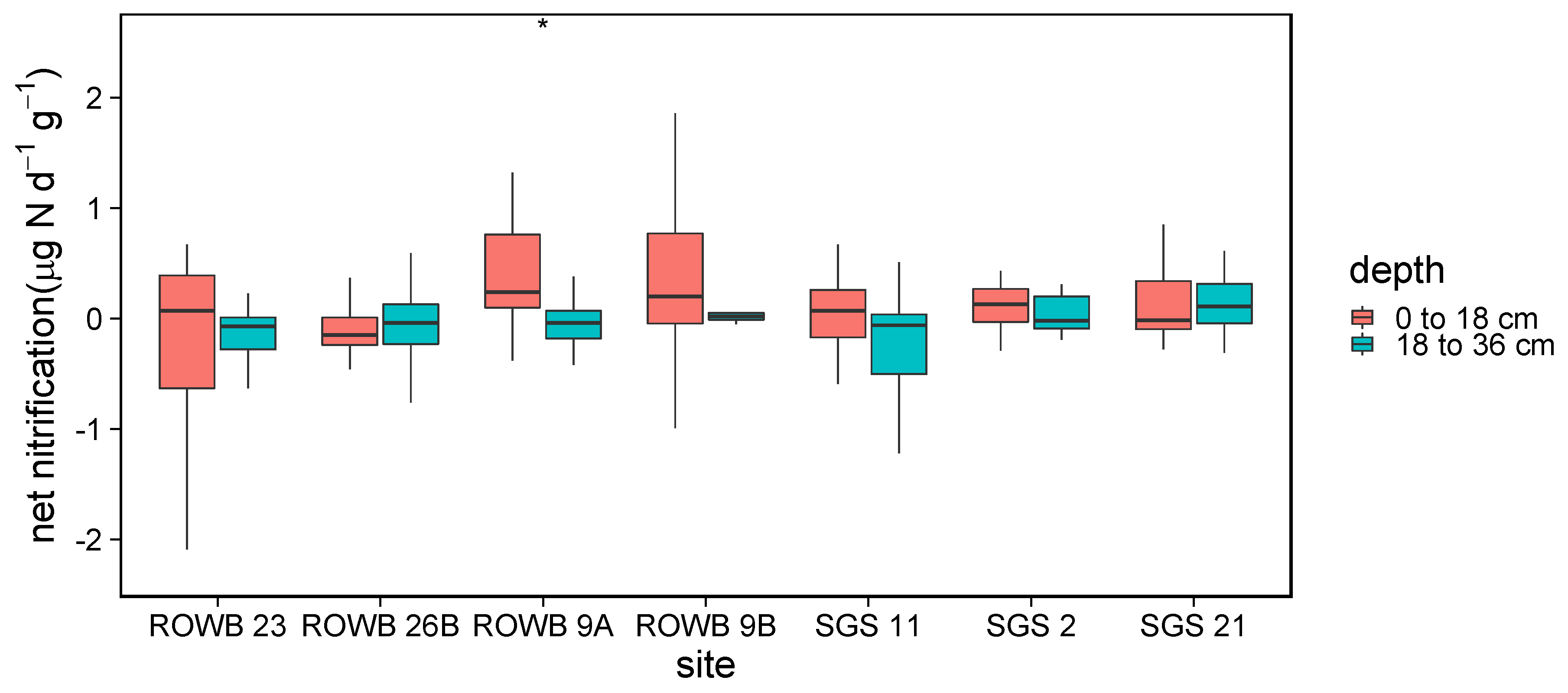
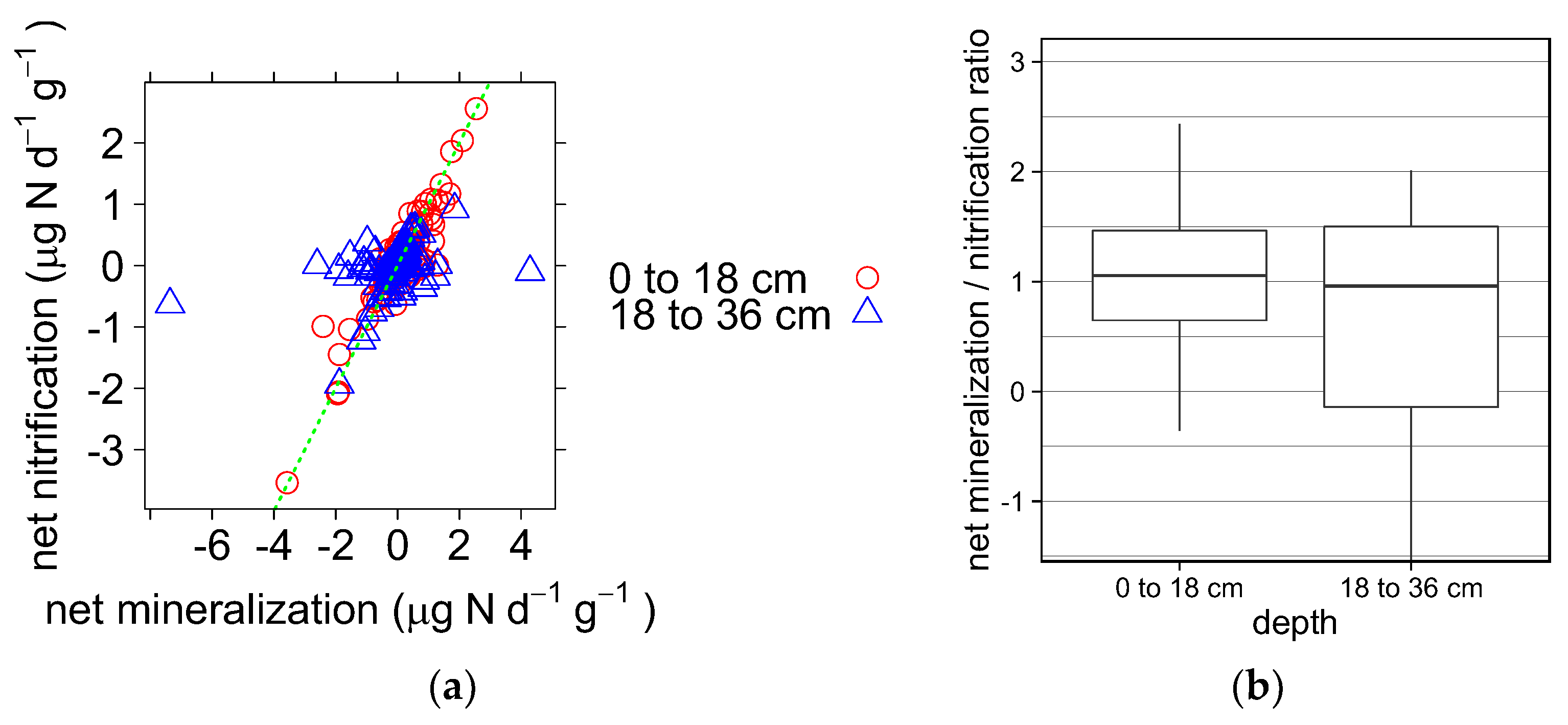
3.1.2. Soil Mineralization and Nitrification
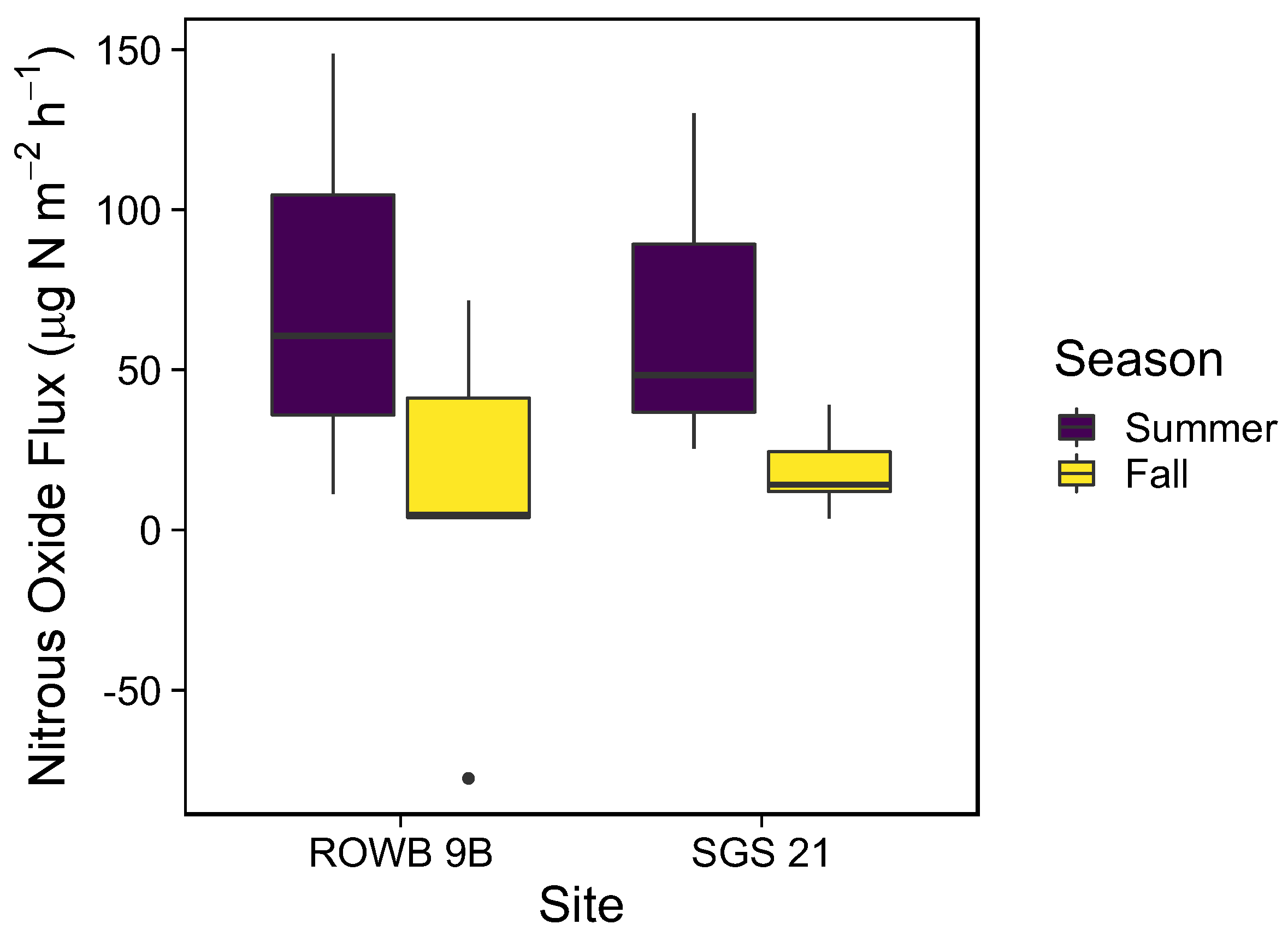
3.2. Soil Gas Emissions
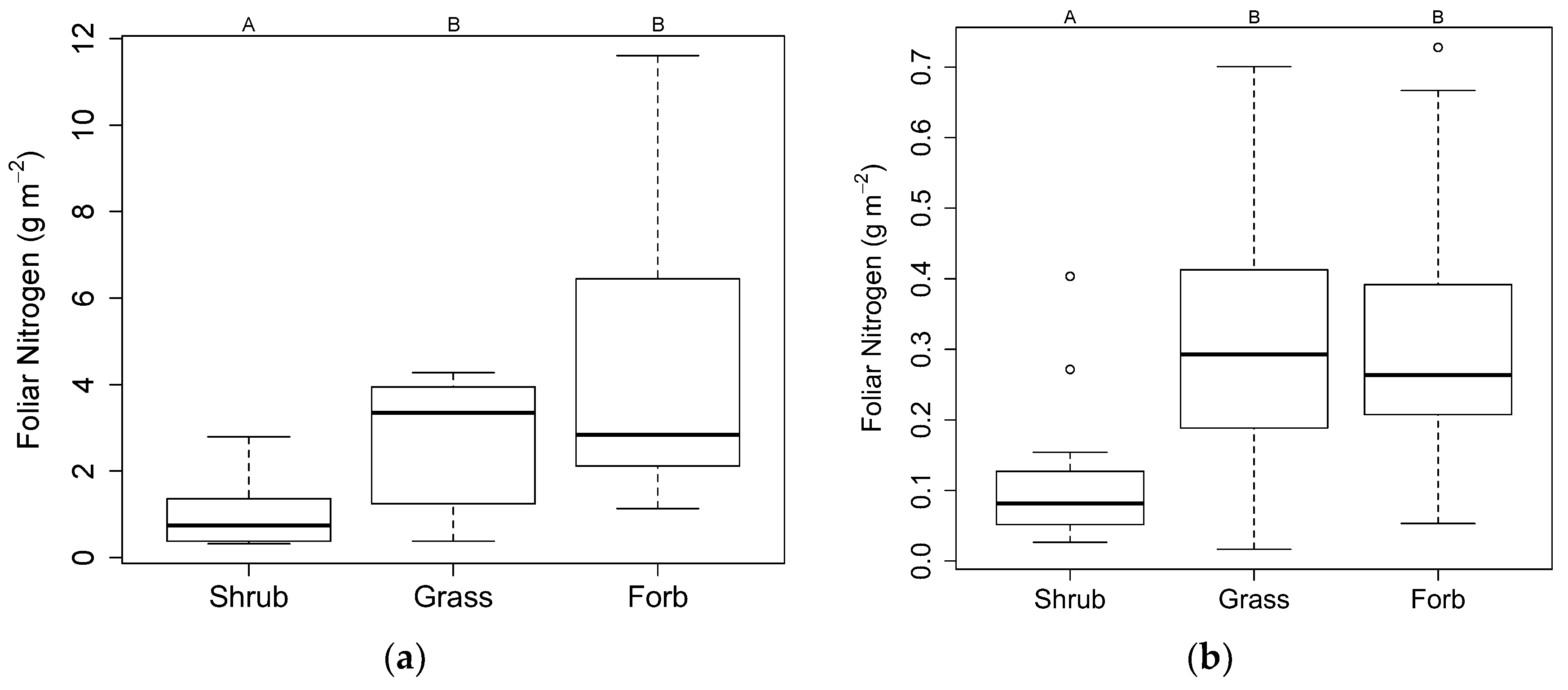
3.3. Plant Uptake
4. Discussion
4.1. Soil
4.1.1. Soil Characterization
4.1.2. Soil Mineralization and Nitrification
4.2. Soil Gas Emissions
4.3. Plant Uptake
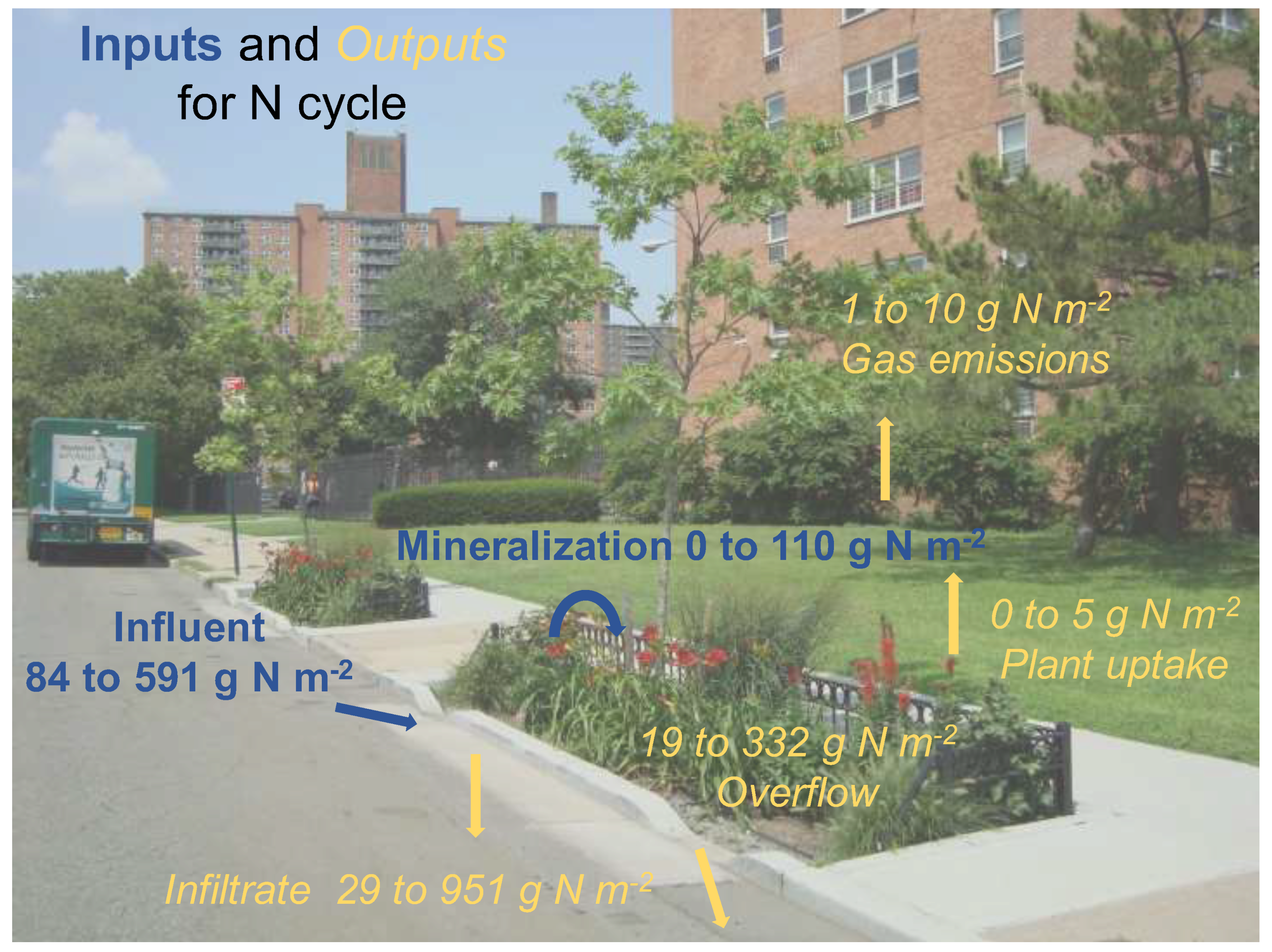
4.4. Nitrogen Mass Balance
4.5. Limitations
4.6. Implications
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Howarth, R.W.; Marino, R. Nitrogen as the Limiting Nutrient for Eutrophication in Coastal Marine Ecosystems: Evolving Views over Three Decades. Limnol. Oceanogr. 2006, 51, 364–376. [Google Scholar] [CrossRef]
- Bettez, N.D.; Groffman, P.M. Nitrogen Deposition in and near an Urban Ecosystem. Environ. Sci. Technol. 2013, 47, 6047–6051. [Google Scholar] [CrossRef] [PubMed]
- Clar, M.; Green, R. Design Manual for Use of Bioretention in Stormwater Management; Department of Environmental Resources: Prince George’s County, MD, USA, 1993. [Google Scholar]
- Bratieres, K.; Fletcher, T.D.; Deletic, A.; Zinger, Y. Nutrient and Sediment Removal by Stormwater Biofilters: A Large-Scale Design Optimisation Study. Water Res. 2008, 42, 3930–3940. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.A.; Lawrence, T.J.; Stander, E.K.; Jontos, R.J.; Kaushal, S.S.; Newcomer, T.A.; Grimm, N.B.; Cole Ekberg, M.L. Opportunities and Challenges for Managing Nitrogen in Urban Stormwater: A Review and Synthesis. Ecol. Eng. 2010, 36, 1507–1519. [Google Scholar] [CrossRef]
- Hunt, W.F.; Jarrett, A.R.; Smith, J.T.; Sharkey, L.J. Evaluating Bioretention Hydrology and Nutrient Removal at Three Field Sites in North Carolina. J. Irrig. Drain. Eng. 2006, 132, 600–608. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Davis, A.P. Urban Stormwater Runoff Nitrogen Composition and Fate in Bioretention Systems. Environ. Sci. Technol. 2014, 48, 3403–3410. [Google Scholar] [CrossRef] [PubMed]
- Passeport, E.; Hunt, W.F. Asphalt Parking Lot Runoff Nutrient Characterization for Eight Sites in North Carolina, USA. J. Hydrol. Eng. 2009, 14, 352–361. [Google Scholar] [CrossRef]
- Peterson, I.J.; Igielski, S.; Davis, A.P. Enhanced Denitrification in Bioretention Using Woodchips as an Organic Carbon Source. J. Sustain. Water Built Environ. 2015, 9, 1–9. [Google Scholar] [CrossRef]
- Davis, A.P.; Shokouhian, M.; Sharma, H.; Minami, C. Water Quality Improvement through Bioretention Media: Nitrogen and Phosphorus Removal. Water Environ. Res. 2006, 78, 284–293. [Google Scholar] [CrossRef] [PubMed]
- McNett, J.K.; Hunt, W.F.; Davis, A.P. Influent Pollutant Concentrations as Predictors of Effluent Pollutant Concentrations for Mid-Atlantic Bioretention. J. Environ. Eng. 2011, 137, 790–799. [Google Scholar] [CrossRef]
- Payne, E.G.I.; Fletcher, T.D.; Cook, P.L.M.; Deletic, A.; Hatt, B.E. Processes and Drivers of Nitrogen Removal in Stormwater Biofiltration. Crit. Rev. Environ. Sci. Technol. 2014, 44, 796–846. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen Mineralization: Challenges of a Changing Paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Robertson, P.G.; Wedin, D.; Groffman, P.M.; Blair, J.M.; Holland, E.A.; Nadelhoffer, K.J.; Harris, D. Soil Carbon and Nitrogen Availability: Nitrogen Mineralization, Nitrification, and Soil Respiration Potentials. In Standard Soil Methods for Long-Term Ecological Research; Robertson, G.P., Bledsoe, C.S., Coleman, D.C., Sollins, P., Eds.; Oxford University Press: New York, NY, USA, 1999; pp. 258–271. [Google Scholar]
- Groffman, P.M.; Boulware, N.J.; Zipperer, W.C.; Pouyat, R.V.; Band, L.E.; Colosimo, M.F. Soil Nitrogen Cycle Processes in Urban Riparian Zones. Environ. Sci. Technol. 2002, 36, 4547–4552. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.P.; Groffman, P.M. Chapter 14—Nitrogen Transformations. In Soil Microbiology Ecology and Biochemistry; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 421–446. [Google Scholar]
- Lucas, W.C.; Greenway, M. Hydraulic Response and Nitrogen Retention in Bioretention Mesocosms with Regulated Outlets: Part II—Nitrogen Retention. Water Environ. Res. 2011, 83. [Google Scholar] [CrossRef]
- Shetty, N.H.; Culligan, P.J.; Mailloux, B.; McGillis, W.R.; Do, H.Y. Bioretention Infrastructure to Manage the Nutrient Runoff from Coastal Cities. In Geo-Chicago 2016 GSP 273; ASCE: Reston, VA, USA, 2016; pp. 402–411. [Google Scholar]
- Brown, R.A.; Birgand, F.; Hunt, W.F. Analysis of Consecutive Events for Nutrient and Sediment Treatment in Field-Monitored Bioretention Cells. Water Air Soil Pollut. 2013, 224, 1581. [Google Scholar] [CrossRef]
- Lucas, W.C.; Greenway, M. Nutrient Retention in Vegetated and Nonvegetated Bioretention Mesocosms. J. Irrig. Drain. Eng. 2008, 134, 613–623. [Google Scholar] [CrossRef]
- McPhillips, L.; Goodale, C.; Walter, M.T. Nutrient Leaching and Greenhouse Gas Emissions in Grassed Detention and Bioretention Stormwater Basins. J. Sustain. Water Built Environ. 2018, 4, 1–10. [Google Scholar] [CrossRef]
- Davidson, E.A.; Keller, M.; Erickson, H.E.; Verchot, L.V.; Veldkamp, E. Testing a Conceptual Model of Soil Emissions of Nitrous and Nitric Oxides. Bioscience 2000, 50, 667. [Google Scholar] [CrossRef] [Green Version]
- Templer, P.H.; Pinder, R.W.; Goodale, C.L. Effects of Nitrogen Deposition on Greenhouse-Gas Fluxes for Forests and Grasslands of North America. Front. Ecol. Environ. 2012, 10, 547–553. [Google Scholar] [CrossRef]
- Grover, S.P.P.; Cohan, A.; Chan, H.S.; Livesley, S.J.; Beringer, J.; Daly, E. Occasional Large Emissions of Nitrous Oxide and Methane Observed in Stormwater Biofiltration Systems. Sci. Total Environ. 2013, 465, 64–71. [Google Scholar] [CrossRef] [PubMed]
- McPhillips, L.E.; Groffman, P.M.; Schneider, R.L.; Walter, M.T. Nutrient Cycling in Grassed Roadside Ditches and Lawns in a Suburban Watershed. J. Environ. Qual. 2016, 45, 1901. [Google Scholar] [CrossRef] [PubMed]
- Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. Greenhouse Gas Emissions from Soils—A Review. Chem. Erde Geochem. 2016, 76, 327–352. [Google Scholar] [CrossRef]
- Livesley, S.J.; Dougherty, B.J.; Smith, A.J.; Navaud, D.; Wylie, L.J.; Arndt, S.K. Soil-Atmosphere Exchange of Carbon Dioxide, Methane and Nitrous Oxide in Urban Garden Systems: Impact of Irrigation, Fertiliser and Mulch. Urban Ecosyst. 2010, 13, 273–293. [Google Scholar] [CrossRef]
- Townsend-Small, A.; Czimczik, C.I. Carbon Sequestration and Greenhouse Gas Emissions in Urban Turf. Geophys. Res. Lett. 2010, 37, 1–5. [Google Scholar] [CrossRef]
- Townsend-Small, A.; Pataki, D.E.; Czimczik, C.I.; Tyler, S.C. Nitrous Oxide Emissions and Isotopic Composition in Urban and Agricultural Systems in Southern California. J. Geophys. Res. 2011, 116, G01013. [Google Scholar] [CrossRef]
- Goatley, M.; Hensler, K. Urban Nutrient Management Handbook; Virginia Cooperative Extension: Petersburg, VA, USA, 2011. [Google Scholar]
- Roy-Poirier, A.; Champagne, P.; Filion, Y. Review of Bioretention System Research and Design: Past, Present, and Future. J. Environ. Eng. 2010, 136, 878–889. [Google Scholar] [CrossRef]
- Payne, E.; Fletcher, T.D.; Russell, D.G.; Grace, M.R.; Cavagnaro, T.R.; Evrard, V.; Deletic, A.; Hatt, B.E.; Cook, P.L.M. Temporary Storage or Permanent Removal? The Division of Nitrogen between Biotic Assimilation and Denitrification in Stormwater Biofiltration Systems. PLoS ONE 2014, 9, e90890. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, S.; Borst, M.; Stander, E.K. Factorial Study of Rain Garden Design for Nitrogen Removal. J. Irrig. Drain. Eng. 2014, 140, 04013016. [Google Scholar] [CrossRef]
- Shetty, N.H. New York City’s Green Infrastructure: Impacts on Nutrient Cycling and Improvements in Performance. Ph.D. Thesis, Columbia University, New York, NY, USA, 2018. [Google Scholar]
- ASTM. Standard Practice for Soil Exploration and Sampling by Auger Borings D1452/D1452M—16; ASTM: West Conshohocken, PA, USA, 2016; pp. 3–9. [Google Scholar]
- Raison, R.J.; Connel, M.J.; Khanna, P.K. Methodology for Studying Fluxes of Soil Mineral-N in Situ. Soil Biol. Biochem. 1987, 19, 521–530. [Google Scholar] [CrossRef]
- Hart, S.C.; Stark, J.M.; Davidson, E.A.; Firestone, M.K. Nitrogen Mineralization, Immobilization, and Nitrification. In Methods of Soil Analysis, Part 2. Microbiological and Biochemical Properties; Weaver, R.W., Angle, J.S., Bottomley, P.J., Bezdiek, D.F., Smith, M.S., Tabatabai, M.A., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 985–1018. [Google Scholar]
- Klute, A. Methods of Soil Analysis: Part 1—Physical and Mineralogical Methods, 2nd ed.; Soil Science Society of America: Madison, WI, USA, 1986; Volume 9. [Google Scholar]
- Holmes, R.M.; Aminot, A.; Kerouel, R.; Hooker, B.A.; Peterson, B.J. A Simple and Precise Method for Measuring Ammonium in Marine and Freshwater Ecosystems. Can. J. Fish. Aquat. Sci. 1999, 56, 1801–1808. [Google Scholar] [CrossRef]
- US EPA. Method 9056A Determination of Inorganic Anions by Ion Chromatography. 2007; No. February; pp. 1–19. [Google Scholar]
- Pihlatie, M.K.; Christiansen, J.R.; Aaltonen, H.; Korhonen, J.F.J.; Nordbo, A.; Rasilo, T.; Benanti, G.; Giebels, M.; Helmy, M.; Sheehy, J.; et al. Comparison of Static Chambers to Measure CH4 Emissions from Soils. Agric. For. Meteorol. 2013, 171–172, 124–136. [Google Scholar] [CrossRef]
- Norton, R.A.; Harrison, J.A.; Kent Keller, C.; Moffett, K.B. Effects of Storm Size and Frequency on Nitrogen Retention, Denitrification, and N2O Production in Bioretention Swale Mesocosms. Biogeochemistry 2017, 134, 353–370. [Google Scholar] [CrossRef]
- Sonnentag, O.; Talbot, J.; Chen, J.M.; Roulet, N.T. Using Direct and Indirect Measurements of Leaf Area Index to Characterize the Shrub Canopy in an Ombrotrophic Peatland. Agric. For. Meteorol. 2007, 144, 200–212. [Google Scholar] [CrossRef]
- Trammell, T.L.; Ralston, H.A.; Scroggins, S.A.; Carreiro, M.M. Foliar Production and Decomposition Rates in Urban Forests Invaded by the Exotic Invasive Shrub, Lonicera Maackii. Biol. Invasions 2012, 14, 529–545. [Google Scholar] [CrossRef]
- Xu, C.Y.; Griffin, K.L.; Schuster, W.S.F. Leaf Phenology and Seasonal Variation of Photosynthesis of Invasive Berberis Thunbergii (Japanese Barberry) and Two Co-Occurring Native Understory Shrubs in a Northeastern United States Deciduous Forest. Oecologia 2007, 154, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Madakadze, I.C.; Stewart, K.; Peterson, P.R.; Coulman, B.E.; Samson, R.; Smith, D.L. Light Interception, Use-Efficiency and Energy Yield of Switchgrass (Panicum virgatum L.) Grown in a Short Season Area. Biomass Bioenergy 1998, 15, 475–482. [Google Scholar] [CrossRef]
- Zeri, M.; Anderson-Teixeira, K.; Hickman, G.; Masters, M.; DeLucia, E.; Bernacchi, C.J. Carbon Exchange by Establishing Biofuel Crops in Central Illinois. Agric. Ecosyst. Environ. 2011, 144, 319–329. [Google Scholar] [CrossRef]
- Kiniry, J.R.; Anderson, L.C.; Johnson, M.V.; Behrman, K.D.; Brakie, M.; Burner, D.; Hawkes, C. Perennial Biomass Grasses and the Mason–Dixon Line: Comparative Productivity across Latitudes in the Southern Great Plains. Bioenergy Res. 2013, 6, 276–291. [Google Scholar] [CrossRef]
- Mitchell, R.B.; Moser, L.E.; Moore, K.J.; Redfearn, D.D. Tiller Demographics and Leaf Area Index of Four Perennial Pasture Grasses. Agron. J. 1998, 90, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Redfearn, D.D.; Moore, K.J.; Vogel, K.P.; Waller, S.S.; Mitchell, R.B. Canopy Architecture and Morphology of Switchgrass Populations Differing in Forage Yield. Agron. J. 1997, 89, 262–269. [Google Scholar] [CrossRef]
- Boyd, N.S.; Gordon, R.; Martin, R.C. Relationship between Leaf Area Index and Ground Cover in Potato under Different Management Conditions. Potato Res. 2002, 45, 117–129. [Google Scholar] [CrossRef]
- Gordon, R.; Brown, D.M.; Dixon, M.A. Estimating Potato Leaf Area Index for Specific Cultivars. Potato Res. 1997, 40, 251–266. [Google Scholar] [CrossRef]
- Helsel, D.R.; Hirsch, R.M. Statistical Methods in Water Resources; Elsevier Inc.: Amsterdam, The Netherlands, 2002; Volume 4. [Google Scholar]
- Chapin, F.S.; Matson, P.A.; Mooney, H.A. Principles of Terrestrial Ecosystem Ecology; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bernot, M.J.; Dodds, W.K. Nitrogen Retention, Removal, and Saturation in Lotic Ecosystems. Ecosystems 2005, 8, 442–453. [Google Scholar] [CrossRef]
- Liu, J.; Sample, D.J.; Owen, J.S.; Li, J.; Evanylo, G. Assessment of Selected Bioretention Blends for Nutrient Retention Using Mesocosm Experiments. J. Environ. Qual. 2014, 43, 1754. [Google Scholar] [CrossRef] [PubMed]
- Sanaullah, M.; Chabbi, A.; Leifeld, J.; Bardoux, G.; Billou, D.; Rumpel, C. Decomposition and Stabilization of Root Litter in Top- and Subsoil Horizons: What Is the Difference? Plant Soil 2011, 338, 127–141. [Google Scholar] [CrossRef]
- Dungait, J.A.J.; Hopkins, D.W.; Gregory, A.S.; Whitmore, A.P. Soil Organic Matter Turnover Is Governed by Accessibility Not Recalcitrance. Glob. Chang. Biol. 2012, 18, 1781–1796. [Google Scholar] [CrossRef]
- Zhang, Z.; Rengel, Z.; Liaghati, T.; Antoniette, T.; Meney, K. Influence of Plant Species and Submerged Zone with Carbon Addition on Nutrient Removal in Stormwater Biofilter. Ecol. Eng. 2011, 37, 1833–1841. [Google Scholar] [CrossRef]
- Elliott, S.; Meyer, M.H.; Sands, G.R.; Horgan, B. Water Quality Characteristics of Three Rain Gardens Located Within the Twin Cities Metropolitan Area, Minnesota. Cities Environ. 2011, 4, 1–15. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Matson, P. Nitrogen Transformations in a Range of Tropical Forest Soils. Soil Biol. Biochem. 1988, 20, 361–367. [Google Scholar] [CrossRef]
- McNulty, S.G.; Aber, J.D.; Mclellan, T.M.; Katt, S.M. Nitrogen Cycling in High Elevation Forests of the Northeastern US in Relation to Nitrogen Deposition. Ambio 1990, 19, 38–40. [Google Scholar]
- Auyeung, D.S.N.; Suseela, V.; Dukes, J.S. Warming and Drought Reduce Temperature Sensitivity of Nitrogen Transformations. Glob. Chang. Biol. 2013, 19, 662–676. [Google Scholar] [CrossRef] [PubMed]
- Nadelhoffer, K.J.; Aber, J.D.; Melillo, J.M. Seasonal Patterns of Ammonium and Nitrate Uptake in Nine Temperate Forest Ecosystems. Plant Soil 1984, 80, 321–335. [Google Scholar] [CrossRef]
- Kastovska, E.; Edwards, K.; Picek, T.; Santruckova, H. A Larger Investment into Exudation by Competitive versus Conservative Plants Is Connected to More Coupled Plant-Microbe N Cycling. Biogeochemistry 2015, 122, 47–59. [Google Scholar] [CrossRef]
- Hatt, B.E.; Fletcher, T.D.; Deletic, A. Hydrologic and Pollutant Removal Performance of Stormwater Biofiltration Systems at the Field Scale. J. Hydrol. 2009, 365, 310–321. [Google Scholar] [CrossRef]
- Shrestha, P.; Hurley, S.E.; Carol Adair, E. Soil Media CO2 and N2O Fluxes Dynamics from Sand-Based Roadside Bioretention Systems. Water 2018, 10, 185. [Google Scholar] [CrossRef]
- Turk, R.P.; Kraus, H.T.; Hunt, W.F.; Carmen, N.B.; Bilderback, T.E. Nutrient Sequestration by Vegetation in Bioretention Cells Receiving High Nutrient Loads. J. Environ. Eng. 2017, 143, 06016009. [Google Scholar] [CrossRef]
- Payne, E.G.I.; Pham, T.; Cook, P.L.M.; Deletic, A.; Hatt, B.E.; Fletcher, T.D. Inside Story of Gas Processes within Stormwater Biofilters: Does Greenhouse Gas Production Tarnish the Benefits of Nitrogen Removal? Environ. Sci. Technol. 2017, 51, 3703–3713. [Google Scholar] [CrossRef] [PubMed]
- Morse, N.R.; McPhillips, L.E.; Shapleigh, J.P.; Walter, M.T. The Role of Denitrification in Stormwater Detention Basin Treatment of Nitrogen. Environ. Sci. Technol. 2017, 51, 7928–7935. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Hunt, W.F.; Traver, R.G.; Clar, M. Bioretention Technology: Overview of Current Practice and Future Needs. J. Environ. Eng. 2009, 135, 109–117. [Google Scholar] [CrossRef]
- Hunt, W.F.; Davis, A.P.; Traver, R.G. Meeting Hydrologic and Water Quality Goals through Targeted Bioretention Design. J. Environ. Eng. 2012, 138, 698–707. [Google Scholar] [CrossRef]
- Woods End Research Laboratory. Guide to Solvita ® Testing for Compost Maturity Index; Woods End Research Laboratory: Mount Vernon, ME, USA, 2002; Volume 3. [Google Scholar]
- Thompson, W.H.; Leege, P.B.; Millner, P.D.; Watson, M.E. Test Methods for the Examination of Composting and Compost (TMECC); Holbrook: New York, NY, USA, 2001. [Google Scholar]
- Matteson, T.L.; Sullivan, D.M. Stability Evaluation of Mixed Food Waste Composts. Compost Sci. Util. 2006, 14, 170–177. [Google Scholar] [CrossRef]
- Analysis of Bioretention Soil Media for Improved Nitrogen, Phosphorus and Copper Retention; Herrera Environmental Consultants: Seattle, WA, USA, 2015.
- Herms, D.A. Understanding Tree Responses to Abiotic and Biotic Stress Complexes. Arborist News 1998, 7, 21–26. [Google Scholar]
- Levin, L.A.; Mehring, A.S. Optimization of Bioretention Systems through Application of Ecological Theory. Wiley Interdiscip. Rev. Water 2015, 2, 259–270. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioswale | Area (m2) | Soil Characterization | Soil Mineralization and Nitrification | Gas Emissions | Plant Uptake |
|---|---|---|---|---|---|
| ROWB 26B | 9.3 | x | xx | x | |
| ROWB 23 | 7.0 | x | x | x | |
| ROWB 9A | 9.3 | x | x | x | |
| ROWB 9B | 9.3 | x | xx | x | x |
| SGS 21 | 76.2 | x | xx | x | |
| SGS 11 | 94.8 | x | xx | ||
| SGS 2 | 115.2 | x | x |
| Depth | Total N (%) | Total C (%) | C:N ratio | Organic (%) | Sand (%) | Silt (%) | Clay (%) |
|---|---|---|---|---|---|---|---|
| 7.6–22.9 cm (n = 7) | 0.15 ± 0.02 | 2.9 ± 0.5 | 19.4 ± 0.4 | 5.0 ± 0.8 | 80.1 ± 1.2 | 13.4 ± 1.3 | 6.5 ± 0.4 |
| 30.5–45.7 cm (n = 7) | 0.13 ± 0.01 | 2.4 ± 0.3 | 18.7 ± 1.2 | 4.1 ± 0.5 | 78.6 ± 1.9 | 13.5 ± 1.5 | 7.9 ± 0.6 |
| p-value | 0.71 | 0.80 | 0.22 | 0.75 | 0.48 | 0.95 | 0.14 |
| Month of Incubation Period | Net Mineralization (µg N d−1 g−1) | Net Nitrification (µg N d−1 g−1) | ||
|---|---|---|---|---|
| June 2015 | −0.81 | b | 0.01 | ab |
| July 2015 | 0.68 | a | 0.49 | ab |
| August 2015 | −0.11 | ab | −0.05 | ab |
| September 2015 | 0.02 | ab | 0.30 | a |
| October 2015 | −0.11 | ab | −0.31 | b |
| November 2015 | 0.10 | ab | 0.03 | ab |
| December 2015 | 0.04 | ab | 0.11 | ab |
| March 2016 | 0.23 | ab | 0.07 | ab |
| April 2016 | 0.28 | ab | 0.12 | ab |
| May 2016 | −0.05 | ab | 0.02 | ab |
| June 2016 | −0.72 | b | −0.50 | b |
| July 2016 | 0.19 | ab | 0.12 | a |
| August 2016 | 0.20 | ab | 0.29 | ab |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shetty, N.; Hu, R.; Hoch, J.; Mailloux, B.; Palmer, M.; Menge, D.N.L.; McGuire, K.; McGillis, W.; Culligan, P. Quantifying Urban Bioswale Nitrogen Cycling in the Soil, Gas, and Plant Phases. Water 2018, 10, 1627. https://doi.org/10.3390/w10111627
Shetty N, Hu R, Hoch J, Mailloux B, Palmer M, Menge DNL, McGuire K, McGillis W, Culligan P. Quantifying Urban Bioswale Nitrogen Cycling in the Soil, Gas, and Plant Phases. Water. 2018; 10(11):1627. https://doi.org/10.3390/w10111627
Chicago/Turabian StyleShetty, Nandan, Ranran Hu, Jessica Hoch, Brian Mailloux, Matthew Palmer, Duncan N. L. Menge, Krista McGuire, Wade McGillis, and Patricia Culligan. 2018. "Quantifying Urban Bioswale Nitrogen Cycling in the Soil, Gas, and Plant Phases" Water 10, no. 11: 1627. https://doi.org/10.3390/w10111627