





Abstract
The purpose of this study was to assess insulin-stimulated gene expression in canine skeletal muscle with a particular focus on NPPC, the gene that encodes C-type natriuretic peptide, a key hormonal regulator of cardiometabolic function. Four conscious canines underwent hyperinsulinemic, euglycemic clamp studies. Skeletal muscle biopsy and arterial plasma samples were collected under basal and insulin-stimulated conditions. Bulk RNA sequencing of muscle tissue was performed to identify differentially expressed genes between these 2 steady-state conditions. Our results showed that NPPC was the most highly expressed gene in skeletal muscle in response to insulin infusion, rising 4-fold between basal and insulin-stimulated conditions. In support of our RNA sequencing data, we found that raising the plasma insulin concentration 15-fold above basal elicited a 2-fold (P = 0.0001) increase in arterial plasma concentrations of N-terminal prohormone C-type natriuretic peptide. Our data suggest that insulin may play a role in stimulating secretion of C-type natriuretic peptide by skeletal muscle. In this context, C-type natriuretic peptide may act in a paracrine manner to facilitate muscle–vascular bed crosstalk and potentiate insulin-mediated vasodilation. This could serve to enhance insulin and glucose delivery, particularly in the postprandial absorptive state.
Over the past 4 decades, intense efforts have been dedicated to defining how atrial natriuretic peptide and brain natriuretic peptide regulate cardiac structure, blood pressure, and blood volume. The role C-type natriuretic peptide (CNP) plays in regulating cardiometabolic function remains incompletely understood, however. Given that CNP possesses antifibrotic, antihypertrophic, antithrombotic, anti-atherogenic, anti-inflammatory, and vasorelaxing properties [1, 2], leaders in the field have recently dubbed the hormone “The Heart’s Guardian Angel” [3].
To date, the tissues that express CNP and the stimuli that govern its release have been incompletely described [4, 5]. Previous investigations identified CNP in the brain [6, 7], chondrocytes [8], and endothelial cells [9, 10], and suggest endothelial shear stress [11, 12], growth factors [10, 13], and other natriuretic peptides [14] trigger the hormone’s release. This information was largely obtained from in vitro studies. Far less is known about the factors that induce the expression of NPPC, the gene that encodes proCNP, which is then cleaved to produce equimolar amounts of biologically active CNP and its congener, N-terminal proCNP (NT-proCNP) [15, 16].
Over the past 2 decades, an increasing number of studies have revealed skeletal muscle’s role as a secretory organ, which produces and secretes numerous myokines with autocrine, paracrine, and endocrine functions [17-19]. Emerging literature suggests that these myokines exert an effect via muscle–vascular bed crosstalk. Ouchi et al [20] recently showed that the myokine Follistatin-like 1 (Fstl1) was secreted in association with increased Akt signaling. Further, their research suggested that increases in promoted endothelial cell function and revascularization in mice. Because insulin activates Akt [21], we reasoned that it may influence muscle–vascular bed crosstalk to enhance glucose delivery to muscle by affecting gene expression and secretion of other peptides, including CNP, a peptide hormone known to cause capillary vasodilation. Thus, in the present study we aimed to characterize insulin-stimulated differential gene expression in canine skeletal muscle with a particular focus on NPPC, the gene that encodes CNP.
Materials and Methods
Animal Care and Surgical Procedures
Four conscious dogs (Canis lupus familiaris) of male sex were studied. Dogs were housed in a facility that met the standards of the American Association for the Accreditation of Laboratory Animal Care guidelines. They were fed a 65 to 75 kcal/kg/day diet of canned meat and chow (28% protein, 49% carbohydrate, and 23% fat) and the protocol was approved by the Vanderbilt University Medical Center Animal Care Committee. Approximately 16 days prior to experiments, a silastic sampling catheter was placed in the femoral artery and a laparotomy was performed as previously described to place infusion catheters into the jejunal and splenic veins, which drain into the hepatic portal circulation [22]. Experimental inclusion criteria included a leukocyte count < 18 000/mm3, hematocrit > 36%, good appetite, normal bowel movements, and healthy physical appearance.
Experimental Procedures
The dogs were fasted for 18 hours prior to each experiment. On the morning of study, intravenous angiocatheters were placed in the cephalic and saphenous veins for infusion of human insulin, 20% dextrose, and somatostatin. The distal ends of the intraportal catheters and flow probes were exteriorized from their subcutaneous pockets through incisions made under local anesthesia (2% lidocaine). Dogs rested in a Pavlov harness throughout the study.
Each experiment consisted of a 90-minute resting period, a 30-minute basal sampling period, and 2 infusion periods of 150 minutes each in which insulin (Novo Nordisk A/S, Copenhagen, Denmark) and somatostatin (Bachem, Torrance, CA) were infused in a peripheral vein and glucagon (Eli Lilly, Indianapolis, IN) was infused into the portal vein (with rates as shown on the top of Fig. 1A). Plasma hormone and metabolite samples were collected during the final 30 minutes of each infusion period. Insulin was infused to raise the peripheral circulation plasma insulin concentration 4-fold and 25-fold above basal during the first and second infusion periods, respectively. Intraportal glucagon was infused at a rate to maintain the hormone’s plasma concentration at a basal level throughout the study.
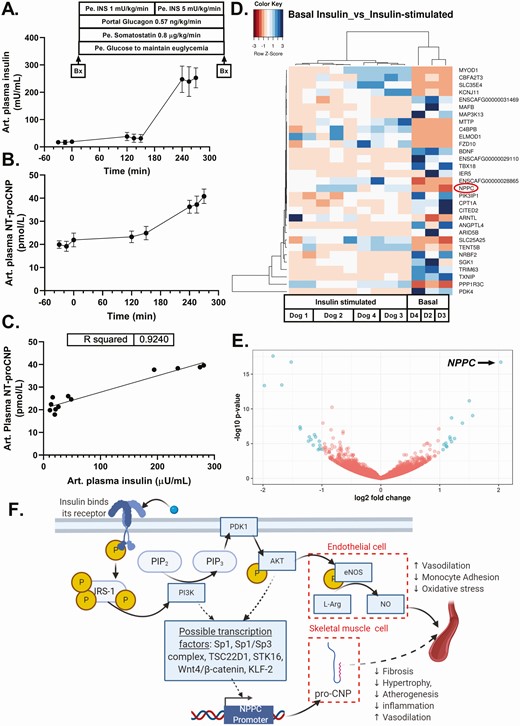
Arterial plasma concentrations (mean ± SD) of (A) insulin and (B) NT-proCNP during a hyperinsulinemic, euglycemic clamp study in 4 dogs. (C) Bivariate relationship between mean arterial plasma levels of insulin and NT-proCNP during each sampling period in each dog. (D) Heatmap depicting differential gene expression in canine skeletal muscle between basal insulin conditions (basal) and insulin-stimulated conditions (insulin-stimulated). Multiple skeletal muscle samples were taken in each dog under insulin-stimulated conditions and 1 sample was taken from each dog during basal conditions. (E) Volcano plot depicting statistical significance (P value) vs magnitude of change (fold change) between basal insulin and insulin-stimulated conditions (red = not significant, blue = significant). NPPC had the greatest magnitude of change, as indicated. (F) Hypothesized mechanism for insulin-mediated CNP transcription and translation. Solid arrows represent known steps in insulin signaling. Hatched arrows represent potential linkages between the insulin signaling cascade and NPPC transcription and proCNP protein synthesis. P represents a phosphorylated signaling intermediate. Abbreviations: Art, arterial; Bx, biopsy. D2, D3, and D4 indicate Dog 2, Dog 3, and Dog 4, respectively.
At 2 points, at the end of the basal period just prior to infusing hormones and immediately following collection of the final plasma hormone and metabolite sample, a <1 cm scalpel incision was made over the biceps femoris muscle and a small muscle biopsy was collected under local anesthesia using sterile technique. Muscle samples were immediately flash frozen in liquid nitrogen and stored at −80 °C.
RNA Sequencing
After completing all canine studies, we dissected 12 frozen muscle samples weighing from 8 to 19 mg each. Total RNA was extracted and purified using Qiagen’s AllPrep kit. Sequencing ready libraries were prepared using Takara SMARTseq v4 for cDNA synthesis kit and addition of sequencing ready adaptors with Nextera XT prep kits respectively. Libraries were sequenced on an Illumina NovaSeq 6000 to an approximate sequencing depth of 40 million total reads.
Read alignment was performed using STAR (v2.7.3a) aligner. The raw read counts were estimated using HTSeq (v0.11.2). Read counts were normalized using DESeq2 to get the normalized counts. Additionally, the aligned reads were used for estimating expression of the genes using cufflinks (v2.2.1). Distribution of mapped reads was performed using RSeQC and RNA-SeQC tools. Analysis was performed using R and additional packages which included: ggplot2, reshape2 and ggrepel, corrplot, gplots and heatmap.2 on normalized counts of all protein coding genes for each sample. Differential expression analysis was performed using DESeq2 (R Bioconductor package, version 3.11). The RNA sequencing data discussed in this publication have been deposited in the National Center for Biotechnology Information’s Gene Expression Omnibus [23] and are accessible through GEO Series accession number GSE167026 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE167026).
Metabolite and Hormone Assays
Arterial plasma glucose levels were assayed using the glucose oxidase reaction (Analox Instruments, Stourbridge, UK). Arterial plasma insulin (Millipore Cat# PI-12K, RRID:AB_2801580, MilliporeSigma, Burlington, MA, USA) and glucagon (Millipore Cat# GL-32K, RRID:AB_2757819, MilliporeSigma) concentrations were measured by radioimmunoassay. Arterial plasma NT-proCNP concentrations were measured using a sandwich ELISA (Biomedica Cat# BI-20812, RRID:AB_2811290, Biomedica, Vienna, Austria).
Statistics
For selecting differentially expressed genes, we used a false-discovery rate adjusted P value of <0.05 and fold change cutoffs of ± 2. A paired t test was used to compare mean arterial plasma concentrations of NT-proCNP between the basal and second insulin-stimulated periods. Simple linear regression was used to analyze the correlation between arterial plasma insulin and NT-proCNP concentrations. Statistical analysis was computed using GraphPad Prism version 8.4.3. Data are summarized as means ± SD unless otherwise indicated. Some or all data generated or analyzed during this study are included in this published article or in the data repositories listed in References.
Results
Arterial Plasma Hormone and Glucose Concentrations
Insulin concentrations rose 2-fold and 15-fold above basal levels during the first and second infusion periods, respectively (Fig. 1A). Glucagon levels remained at basal levels throughout the experiment, with mean plasma concentrations of 53.2 ± 17.0 and 44.9 ± 6.6 pg/mL during the basal and experimental periods, respectively. Plasma glucose concentrations were 91.8 ± 23.7 and 106.3 ± 12.9 mg/dL during the basal and experimental periods, respectively. Mean glucose infusion rates during the sampling period of the first and second infusions were 6.9 ± 3.2 and 19.7 ± 3.0 mg/kg/min, respectively.
RNA Sequencing Data
Bulk RNA sequencing analysis revealed that insulin infusion decreased the expression of 17 genes and increased the expression of 15 genes by more than 2-fold in canine muscle tissue (Fig. 1D and available at [24]). Notably, the gene that was most highly induced (4-fold) in skeletal muscle in response to insulin infusion was NPPC, which encodes for the preproprotein precursor of CNP (Fig. 1E). Other differentially expressed genes included ANGPTL4 and BDNF, which encode for 2 purported myokines in humans [18] (insulin suppressed gene expression to one-quarter and one-third of basal levels of each gene, respectively).
Arterial Plasma NT-proCNP Concentrations
Since CNP is a secreted protein, we evaluated plasma CNP levels under the steady-state conditions of our hyperinsulinemic, euglycemic clamp to determine if the increase in NPPC expression translated into an increase in its secreted form in the circulation. Because bioactive CNP is subject to rapid clearance after secretion [25], we instead assayed plasma levels of its amino-terminal congener, NT-proCNP, which is not rapidly cleared [26]. In support of our RNA sequencing data, raising the plasma insulin concentration 15-fold above basal elicited a 2-fold increase in plasma NT-proCNP (P = 0.0001, Fig. 1B). In addition, plasma concentrations of insulin and NT-proCNP were highly positively correlated (Fig. 1C), suggestive of a strong association between elevations in plasma insulin and increased plasma NT-proCNP.
Discussion
Our canine data show that an increase in the arterial plasma insulin concentration is closely associated with a significant increase in skeletal muscle tissue expression of NPPC, which encodes the preproprotein precursor of CNP. Moreover, our findings suggest increasing levels of plasma insulin may stimulate production of CNP. Taken together, our results imply that skeletal muscle cells secrete CNP as a myokine and that insulin signaling may stimulate this process.
Currently, the transduction pathways that lead to upregulation of NPPC expression are incompletely understood [5]. Previous studies examining the effect of growth factors on NPPC expression identified several potential transcription factors, including Specificity protein 1 (Sp1), an Sp1/Sp3 complex, TSC22 Domain Family Member 1 (TSC22D1), Serine/Threonine Kinase 16 (STK16), Krüppel-like Factor 2 (KLF-2), and the Wnt4/β-catenin pathway [27-31]. Because no study has determined whether CNP production is Akt-dependent (to our knowledge), we think the findings in this study call for further investigation of this possibility. In the vascular endothelium, insulin is known to stimulate vasodilation via activation of endothelial nitric oxide synthase to produce nitric oxide via phosphatidylinositol 3-kinase and Akt activation [32]. Thus, it is possible that an analogous, complementary pathway could exist in skeletal muscle, whereby insulin stimulates the secretion of CNP, leading to muscle–vascular bed crosstalk that potentiates insulin-mediated vasodilation (Fig. 1F). This paracrine effect of CNP would serve to facilitate insulin and glucose delivery, particularly in the absorptive state when insulin levels are relatively high.
Some limitations of this investigation warrant consideration. First, dogs in the present study have a higher percentage muscle mass than humans. Additionally, the fast and oxidative muscle fibers in dogs are relatively fatigue resistant compared to humans. These teleological differences may limit the extent to which our canine data can be extrapolated to humans. Second, the sample size in our study was small, so further studies are needed to validate the link between insulin and CNP. Third, because there was no control group where the plasma insulin was fixed at basal levels our results cannot exclude the possibility that an unmeasured covariate other than insulin may have confounded the relationship between insulin and NT-proCNP levels. Fourth, although the combination of increased NPPC expression in skeletal muscle implies that the rising plasma levels of NT-proCNP during the insulin infusion were caused by increased production of CNP, these data cannot exclude the possibility that the increased NT-proCNP levels were caused by a reduction in NT-proCNP clearance. Fifth, although somatostatin’s effects on the body are generally inhibitory, we cannot rule out the possibility that somatostatin contributed to increased NPPC expression via indirect mechanisms. Sixth, while this in vivo study demonstrates an association between rising plasma insulin concentrations and increases in skeletal muscle NPPC expression and plasma NT-proCNP levels, it cannot exclude the possibility that cell types other than, or in addition to, myocytes may have contributed to the increase in plasma NT-proCNP. Nevertheless, our observation that insulin infusion is associated with an increase in both skeletal muscle tissue expression of NPPC and plasma concentrations of NT-proCNP is unique and should spur further research to elucidate insulin’s role in stimulating CNP synthesis.
Conclusions
Our results suggest insulin plays a role in upregulating CNP gene expression in skeletal muscle and stimulating the secretion of CNP in the circulation. Additional studies are needed to define the mechanism(s) coupling insulin signaling to CNP production and to further characterize which tissues predominate in producing CNP in the body.
Abbreviations
Acknowledgments
The authors thank Amy Nunally, Jamie Adcock, and Donna LeMay for their assistance with animal care.
Financial Support: This work was supported by a gift from Ms. Rob Pierce and the Appleby Foundation. J.M.G. was also supported by the Vanderbilt University Faculty Research Scholars Program, the National Institute of Diabetes and Digestive and Kidney Diseases of the National Institutes of Health under award number K23DK123392, and by a JDRF Career Development Award (5-ECR-2020-950-A-N). The funding sources were not involved in this study’s design; in the collection, analysis, and analysis of data; in the writing of the report; nor in the decision to submit the article for publication. The content is solely the responsibility of the authors and does not necessarily represent the official views of the Appleby Foundation, National Institutes of Health, or the JDRF.
Author Contributions: J.M.G. designed and conducted canine experiments, prepared muscle biopsy samples for RNA sequencing, analyzed the data, and wrote the manuscript. G.K., B.F., M.S.S., P.E.W., and D.C.L. performed canine studies. K.T. and Y.R.S. assayed NT-proCNP. C.S.W. and M.D.T. assisted with RNA sequencing and data analysis. A.D.C. designed canine experiments, analyzed the data, and assisted with writing the manuscript. K.C.C. prepared muscle biopsy samples for RNA sequencing, analyzed the data, and wrote the manuscript. All authors critically reviewed the manuscript, provided input, and gave approval of the final version to be published.
Additional Information
Disclosures: J.M.G. reports consulting fees from InClinica and advisory board fees from Eli Lilly, Medtronic, and Mannkind Corporation. A.D.C. reports grants and consulting fees from Novo Nordisk and is a scientific advisory board member for Biocon. The other authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Data Availability
Some or all data generated or analyzed during this study are included in this published article or in the data repositories listed in References (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE167026).

{kind=link}