
CD146 Delineates an Interfascicular Cell Sub-Population in Tendon That Is Recruited during Injury through Its Ligand Laminin-α4

 , , ,
, , ,
Abstract
:1. Introduction
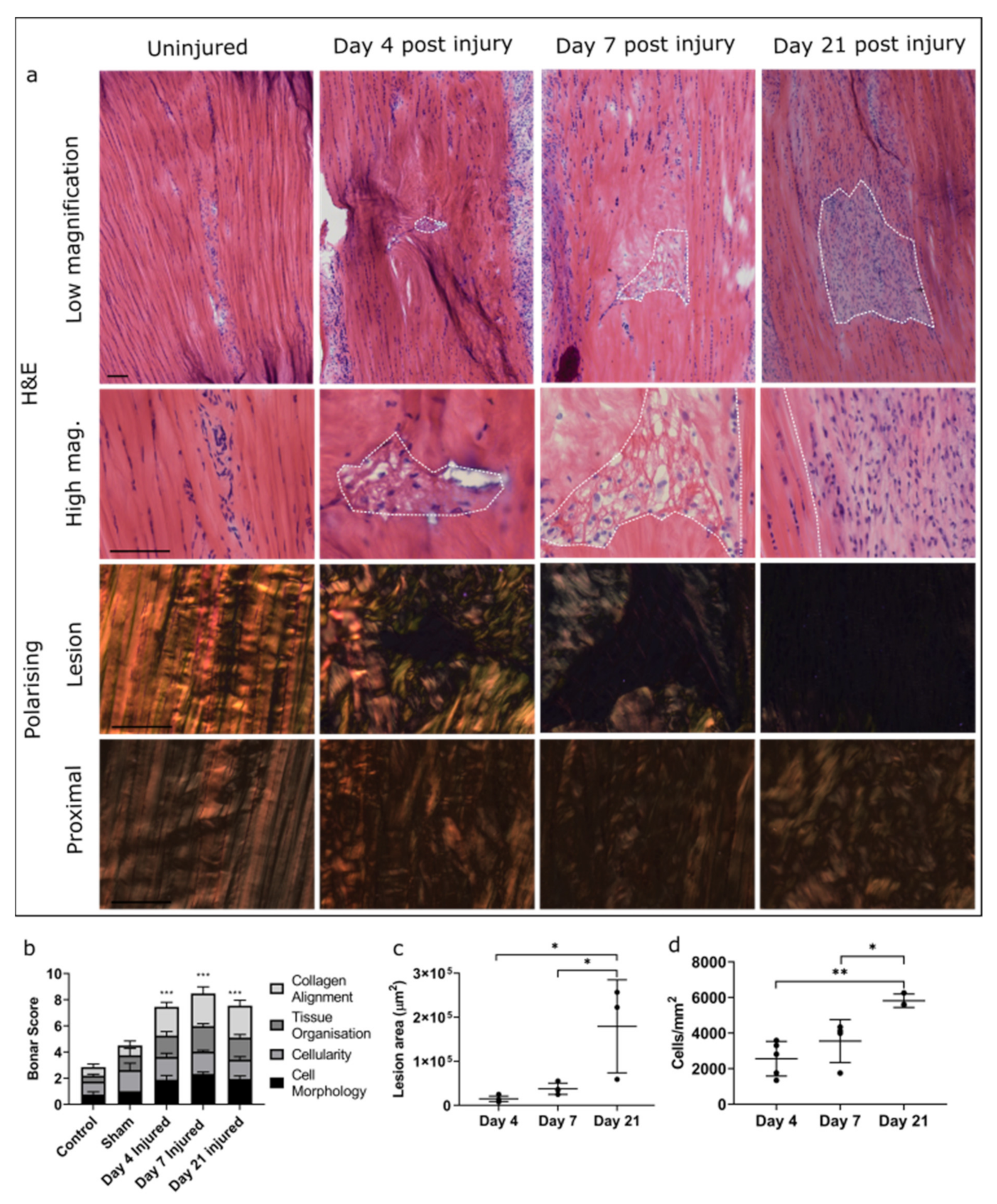
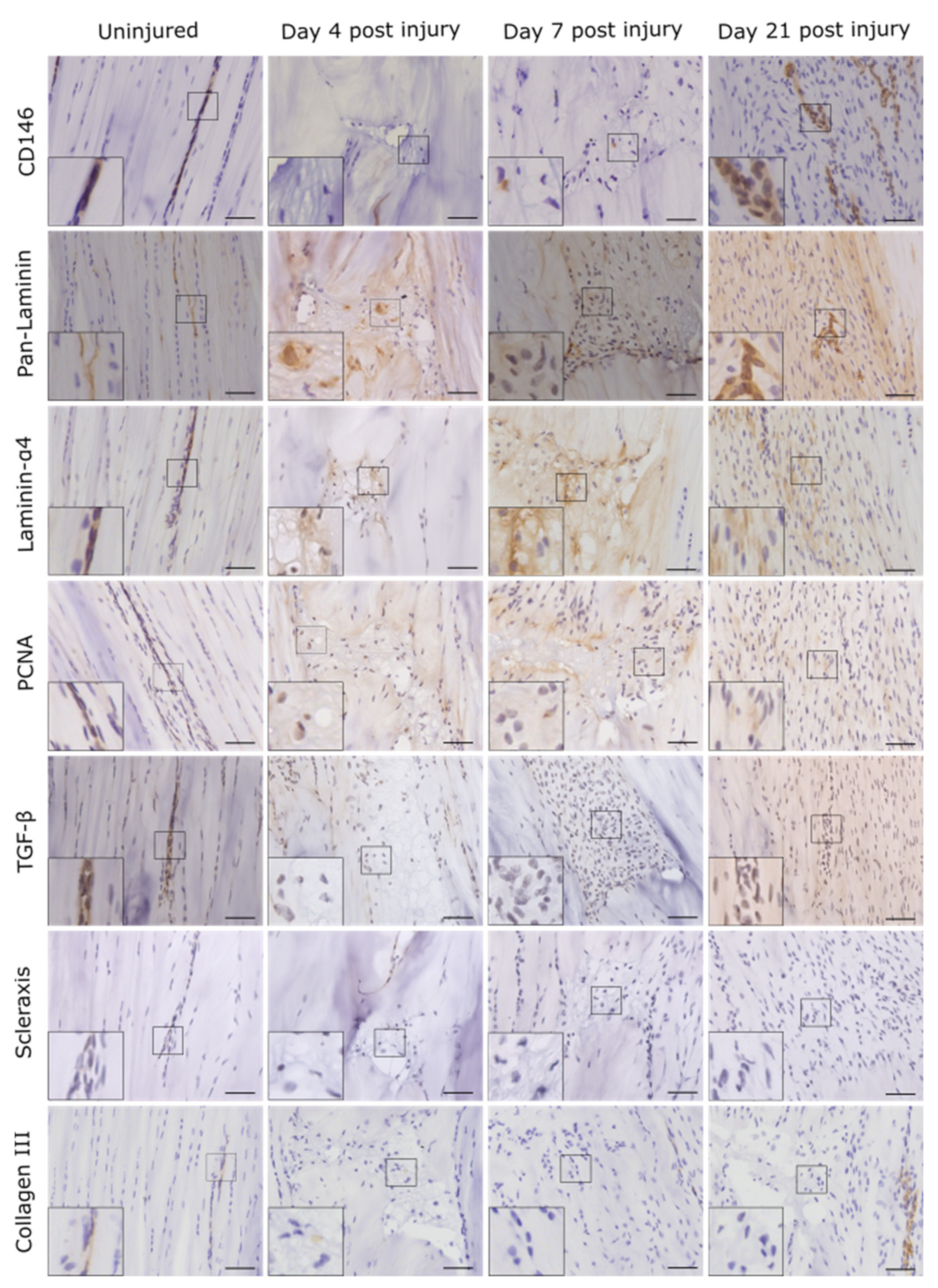
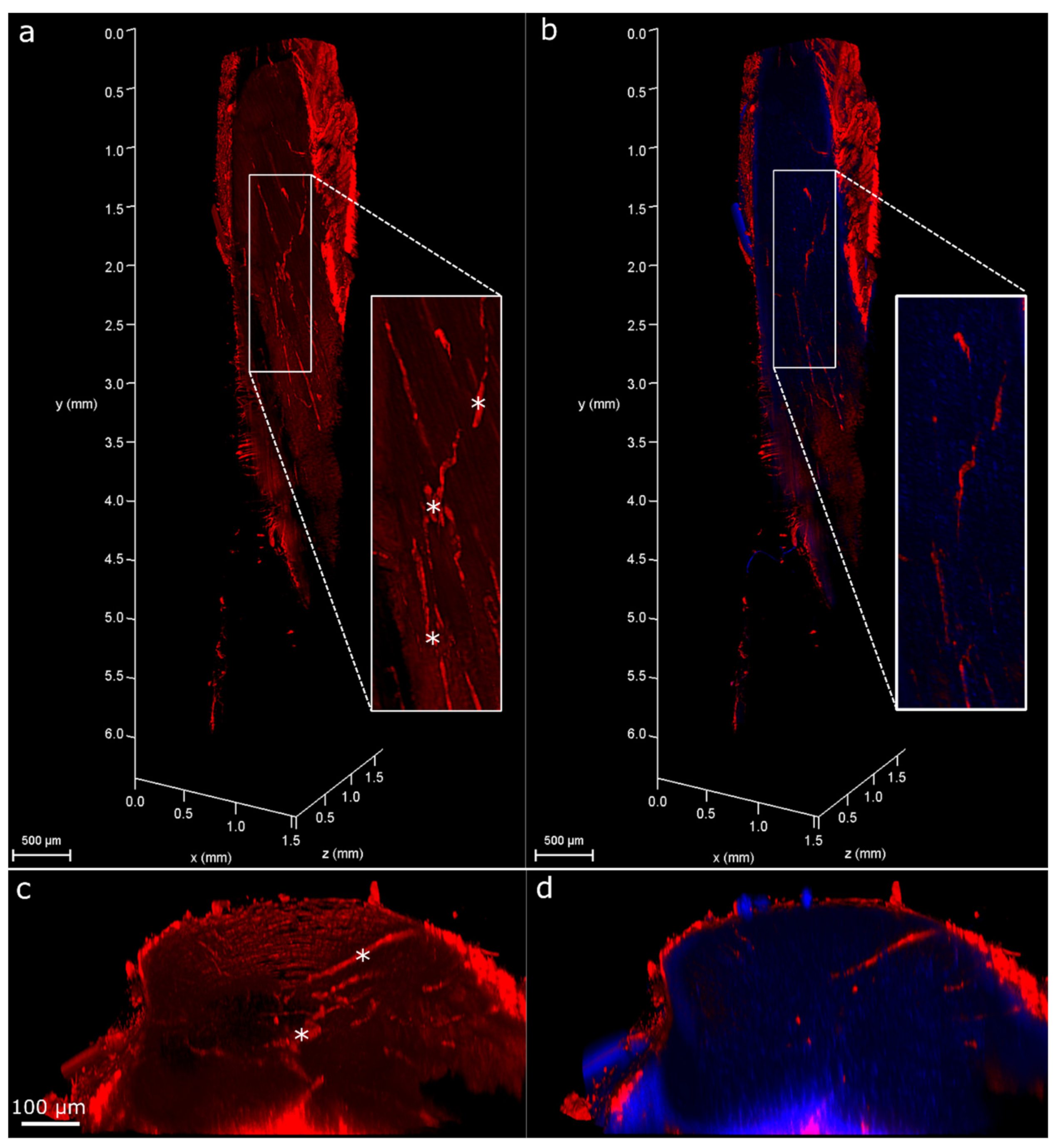
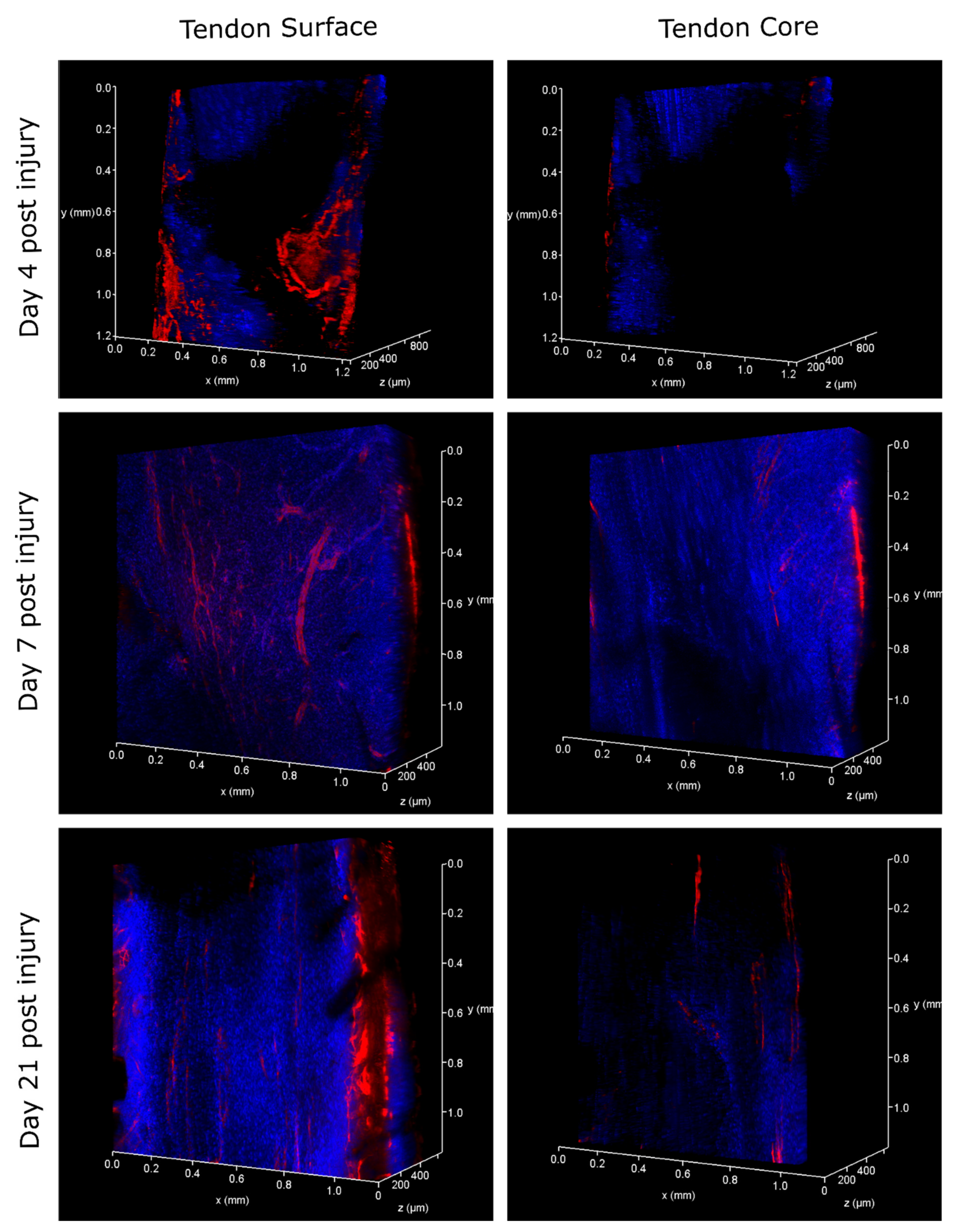
2. Results
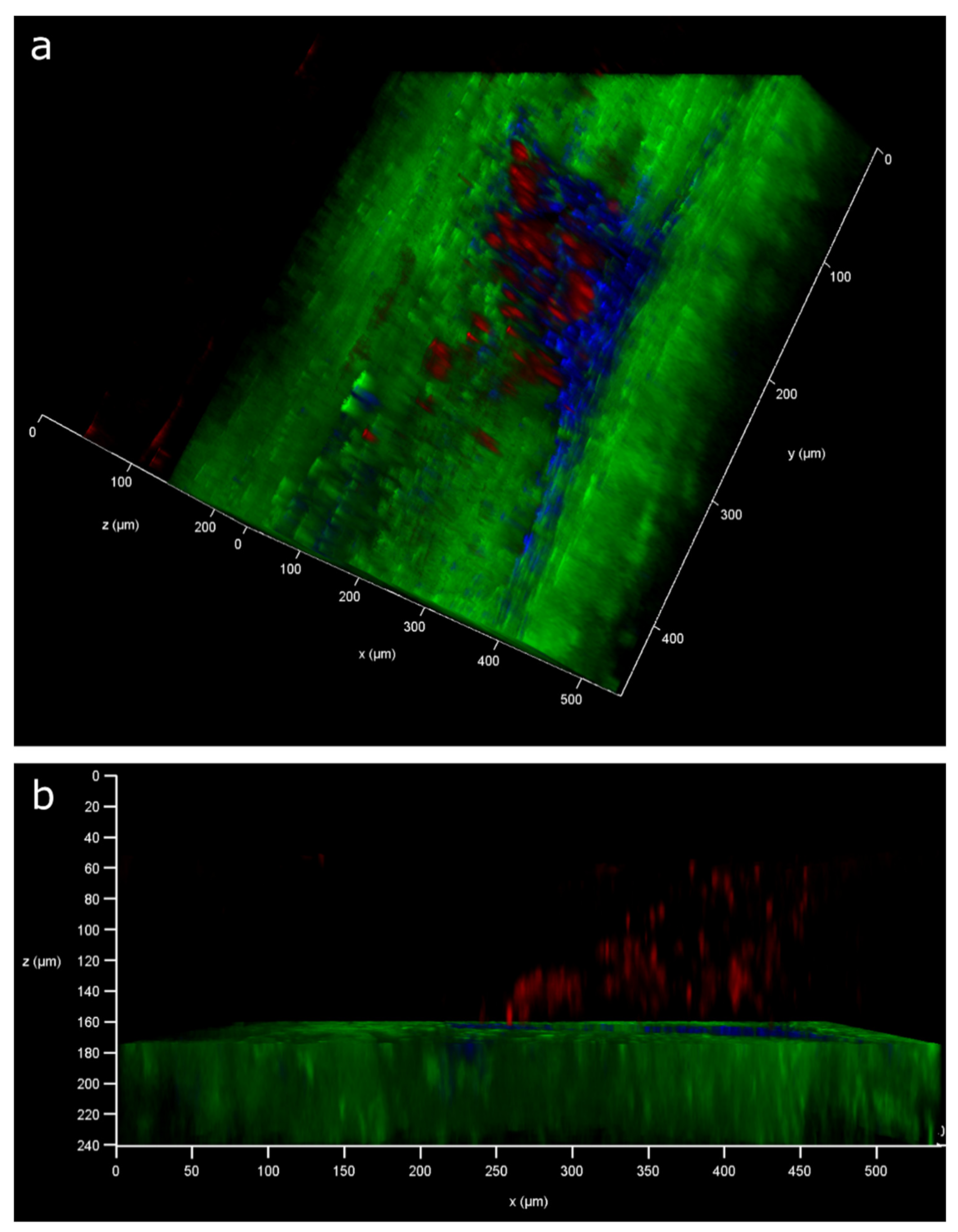
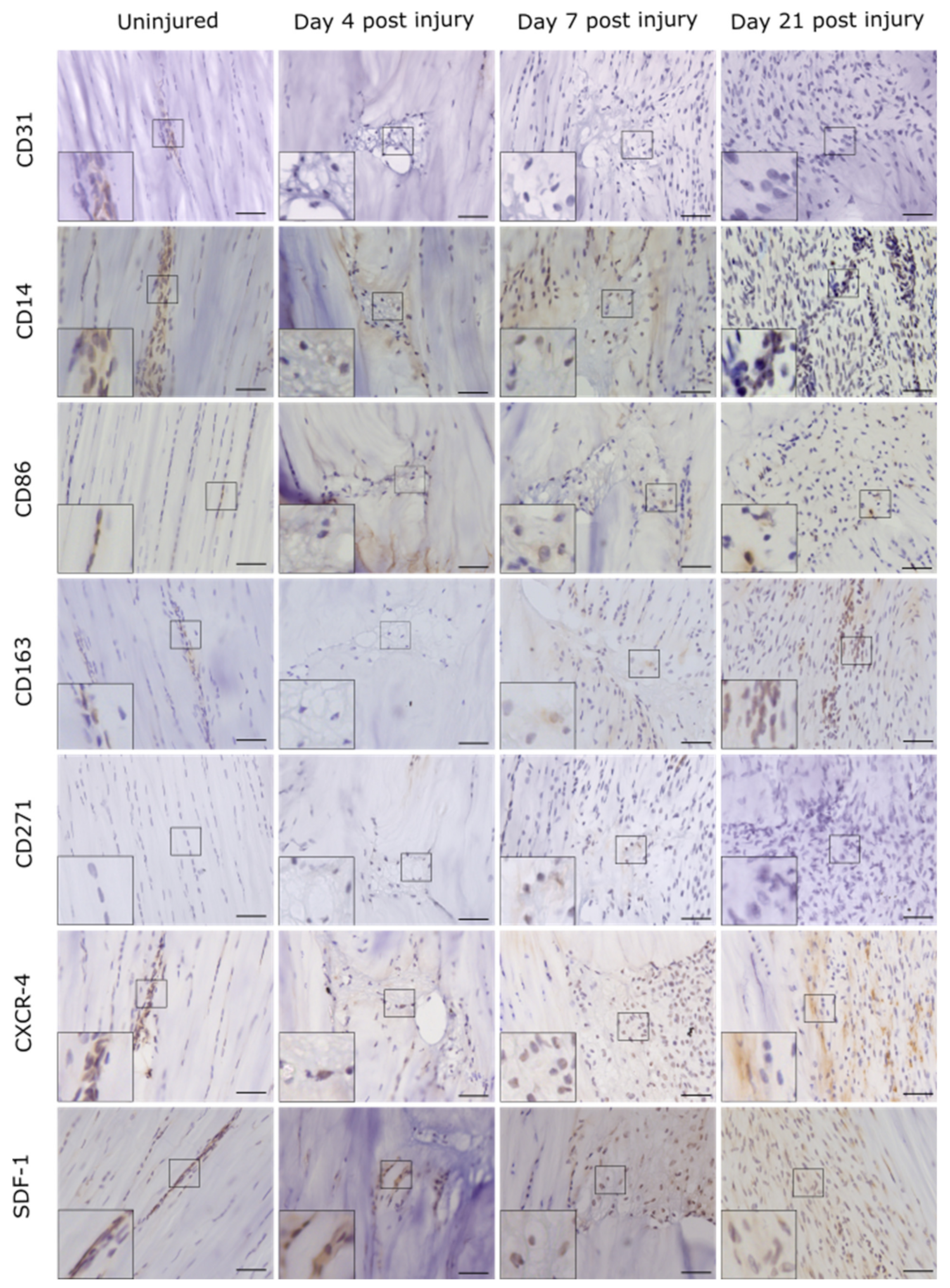
2.1. Injury Study
2.2. Serum RNA-seq
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Tendon Needle Injury
4.3. 2D Analysis of Cell Response to Injury
4.4. Statistical Analysis
4.5. Immunolabelling
4.6. 3D Analysis of Cell Response to Injury
4.7. Confocal Imaging
4.8. RNA Isolation, cDNA Library Preparation, and Small RNA Sequencing (RNA-seq)
4.9. Data Processing
4.10. Pathway Analysis
4.11. qRT-PCR Validation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | Bovine serum albumin |
| DE | Differentially expressed |
| ECM | Extracellular matrix |
| FDR | False discovery rate |
| IFM | Interfascicular matrix |
| IPA | Ingenuity pathway analysis |
| LAMA4 | Laminin-α4 |
| LogFC | Log2 fold change |
| miR | microRNA |
| ncRNA | Non-coding RNA |
| OCT | Optimal cutting temperature |
| PCA | Principle component analysis |
| PCNA | Proliferating Cell Nuclear Antigen |
| PFA | Paraformaldehyde |
| SCX | Scleraxis |
| TBS | Tris-buffered saline |
References
- Thorpe, C.T.; Udeze, C.P.; Birch, H.L.; Clegg, P.D.; Screen, H.R. Specialization of tendon mechanical properties results from interfascicular differences. J. R. Soc. Interface 2012, 9, 3108–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorpe, C.T.; Godinho, M.S.; Riley, G.P.; Birch, H.L.; Clegg, P.D.; Screen, H.R. The interfascicular matrix enables fascicle sliding and recovery in tendon, and behaves more elastically in energy storing tendons. J. Mech. Behav. Biomed. Mater. 2015, 52, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, C.T.; Riley, G.P.; Birch, H.L.; Clegg, P.D.; Screen, H.R. Fascicles and the interfascicular matrix show adaptation for fatigue resistance in energy storing tendons. Acta Biomater. 2016, 42, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, C.T.; Karunaseelan, K.J.; Ng Chieng Hin, J.; Riley, G.P.; Birch, H.L.; Clegg, P.D.; Screen, H.R.C. Distribution of proteins within different compartments of tendon varies according to tendon type. J. Anat. 2016, 229, 450–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godinho, M.S.C.; Thorpe, C.T.; Greenwald, S.E.; Screen, H.R.C. Elastin is localised to the interfascicular matrix of energy storing tendons and becomes increasingly disorganised with ageing. Sci. Rep. 2017, 7, 9713. [Google Scholar] [CrossRef] [PubMed]
- Kohrs, R.T.; Zhao, C.; Sun, Y.-L.; Jay, G.D.; Zhang, L.; Warman, M.L.; An, K.-N.; Amadio, P.C. Tendon fascicle gliding in wild type, heterozygous, and lubricin knockout mice. J. Orthop. Res. 2011, 29, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Godinho, M.S.; Thorpe, C.T.; Greenwald, S.E.; Screen, H.R.C. Elastase treatment of tendon specifically impacts the mechanical properties of the interfascicular matrix. Acta Biomater. 2021, 123, 187–196. [Google Scholar] [CrossRef]
- Choi, H.; Simpson, D.; Wang, D.; Prescott, M.; Pitsillides, A.A.; Dudhia, J.; Clegg, P.D.; Ping, P.; Thorpe, C.T. Heterogeneity of proteome dynamics between connective tissue phases of adult tendon. eLife 2020, 9, e55262. [Google Scholar] [CrossRef]
- Thorpe, C.T.; Peffers, M.J.; Simpson, D.; Halliwell, E.; Screen, H.R.; Clegg, P.D. Anatomical heterogeneity of tendon: Fascicular and interfascicular tendon compartments have distinct proteomic composition. Sci. Rep. 2016, 6, 20455. [Google Scholar] [CrossRef]
- Spiesz, E.M.; Thorpe, C.T.; Chaudhry, S.; Riley, G.P.; Birch, H.L.; Clegg, P.D.; Screen, H.R. Tendon extracellular matrix damage, degradation and inflammation in response to in vitro overload exercise. J. Orthop. Res. 2015, 33, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Marr, N.; Hopkinson, M.; Hibbert, A.P.; Pitsillides, A.A.; Thorpe, C.T. Bimodal whole-mount imaging of tendon using confocal microscopy and X-ray micro-computed tomography. Biol. Proc. Online 2020, 22, 13. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V. Basement membrane proteoglycans: From cellar to ceiling. Nat. Rev. Mol. Cell Biol. 2005, 6, 646–656. [Google Scholar] [CrossRef]
- Sanes, J.R. The basement membrane/basal lamina of skeletal muscle. J. Biol. Chem. 2003, 278, 12601–12604. [Google Scholar] [CrossRef] [Green Version]
- Wragg, J.W.; Finnity, J.P.; Anderson, J.A.; Ferguson, H.J.; Porfiri, E.; Bhatt, R.I.; Murray, P.G.; Heath, V.L.; Bicknell, R. MCAM and LAMA4 are highly enriched in tumor blood vessels of renal cell carcinoma and predict patient outcome. Cancer Res. 2016, 76, 2314–2326. [Google Scholar] [CrossRef] [Green Version]
- Thyboll, J.; Kortesmaa, J.; Cao, R.; Soininen, R.; Wang, L.; Iivanainen, A.; Sorokin, L.; Risling, M.; Cao, Y.; Tryggvason, K. Deletion of the laminin alpha4 chain leads to impaired microvessel maturation. Mol. Cell Biol. 2002, 22, 1194–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaicik, M.K.; Blagajcevic, A.; Ye, H.; Morse, M.C.; Yang, F.; Goddi, A.; Brey, E.M.; Cohen, R.N. The absence of laminin α4 in male mice results in enhanced energy expenditure and increased beige subcutaneous adipose tissue. Endocrinology 2018, 159, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Susek, K.H.; Korpos, E.; Huppert, J.; Wu, C.; Savelyeva, I.; Rosenbauer, F.; Müller-Tidow, C.; Koschmieder, S.; Sorokin, L. Bone marrow laminins influence hematopoietic stem and progenitor cell cycling and homing to the bone marrow. Matrix Biol. 2018, 67, 47–62. [Google Scholar] [CrossRef]
- Flanagan, K.; Fitzgerald, K.; Baker, J.; Regnstrom, K.; Gardai, S.; Bard, F.; Mocci, S.; Seto, P.; You, M.; Larochelle, C.; et al. Laminin-411 is a vascular ligand for MCAM and facilitates TH17 cell entry into the CNS. PLoS ONE 2012, 7, e40443. [Google Scholar] [CrossRef]
- Yin, Z.; Hu, J.-J.; Yang, L.; Zheng, Z.-F.; An, C.-R.; Wu, B.-B.; Zhang, C.; Shen, W.-L.; Liu, H.-H.; Chen, J.-L.; et al. Single-cell analysis reveals a nestin+ tendon stem/progenitor cell population with strong tenogenic potentiality. Sci. Adv. 2016, 2, e1600874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohler, J.; Popov, C.; Klotz, B.; Alberton, P.; Prall, W.C.; Haasters, F.; Muller-Deubert, S.; Ebert, R.; Klein-Hitpass, L.; Jakob, F.; et al. Uncovering the cellular and molecular changes in tendon stem/progenitor cells attributed to tendon aging and degeneration. Aging Cell 2013, 12, 988–999. [Google Scholar] [CrossRef] [Green Version]
- Freedman, B.R.; Fryhofer, G.W.; Salka, N.S.; Raja, H.A.; Hillin, C.D.; Nuss, C.A.; Farber, D.C.; Soslowsky, L.J. Mechanical, histological, and functional properties remain inferior in conservatively treated Achilles tendons in rodents: Long term evaluation. J. Biomech. 2017, 56, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korntner, S.; Kunkel, N.; Lehner, C.; Gehwolf, R.; Wagner, A.; Augat, P.; Stephan, D.; Heu, V.; Bauer, H.-C.; Traweger, A.; et al. A high-glucose diet affects Achilles tendon healing in rats. Sci. Rep. 2017, 7, 780. [Google Scholar] [CrossRef] [PubMed]
- Lake, S.P.; Ansorge, H.L.; Soslowsky, L.J. Animal models of tendinopathy. Disabil. Rehabil. 2008, 30, 1530–1541. [Google Scholar] [CrossRef] [PubMed]
- Foland, J.W.; Trotter, G.W.; Powers, B.E.; Wrigley, R.H.; Smith, F.W. Effect of sodium hyaluronate in collagenase-induced superficial digital flexor tendinitis in horses. Am. J. Vet. Res. 1992, 53, 2371–2376. [Google Scholar]
- Da Silva, F.S.; Abreu, B.J.; Eriksson, B.I.; Ackermann, P.W. Complete mid-portion rupture of the rat achilles tendon leads to remote and time-mismatched changes in uninjured regions. Knee Surg. Sports Traumatol. Arthrosc. 2021, 29, 1990–1999. [Google Scholar] [CrossRef]
- O’Brien, E.J.O.; Shrive, N.G.; Rosvold, J.M.; Thornton, G.M.; Frank, C.B.; Hart, D.A. Tendon mineralization is accelerated bilaterally and creep of contralateral tendons is increased after unilateral needle injury of murine achilles tendons. J. Orthop. Res. 2013, 31, 1520–1528. [Google Scholar] [CrossRef]
- O’Brien, E.J.; Frank, C.B.; Shrive, N.G.; Hallgrímsson, B.; Hart, D.A. Heterotopic mineralization (ossification or calcification) in tendinopathy or following surgical tendon trauma. Int. J. Exp. Pathol. 2012, 93, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.L.; Esa, M.S.; Li, K.H.C.; Krishnan, S.R.G.; Elgallab, G.M.; Pearce, M.S.; Young, D.A.; Birrell, F.N. Osteoporosis, fracture, osteoarthritis & sarcopenia: A systematic review of circulating microRNA association. Bone 2021, 152, 116068. [Google Scholar] [CrossRef]
- Grillari, J.; Mäkitie, R.E.; Kocijan, R.; Haschka, J.; Vázquez, D.C.; Semmelrock, E.; Hackl, M. Circulating miRNAs in bone health and disease. Bone 2021, 145, 115787. [Google Scholar] [CrossRef]
- Plachel, F.; Heuberer, P.; Gehwolf, R.; Frank, J.; Tempfer, H.; Lehner, C.; Weissenbacher, N.; Wagner, A.; Weigl, M.; Moroder, P.; et al. MicroRNA profiling reveals distinct signatures in degenerative rotator cuff pathologies. J. Orthop. Res. 2020, 38, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Kocijan, R.; Muschitz, C.; Geiger, E.; Skalicky, S.; Baierl, A.; Dormann, R.; Plachel, F.; Feichtinger, X.; Heimel, P.; Fahrleitner-Pammer, A.; et al. Circulating microRNA signatures in patients with idiopathic and postmenopausal osteoporosis and fragility fractures. J. Clin. Endocrinol. Metab. 2016, 101, 4125–4134. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Ding, H.; Jiang, H.; Bao, N.; Zhou, L.; Zhao, J. miR-338-5p regulates the viability, proliferation, apoptosis and migration of rheumatoid arthritis fibroblast-like synoviocytes by targeting NFAT5. Cell. Physiol. Biochem. 2018, 49, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, J.; Pan, Z.; Zhang, Y. miR-143-3p regulates cell proliferation and apoptosis by targeting IGF1R and IGFBP5 and regulating the Ras/p38 MAPK signaling pathway in rheumatoid arthritis. Exp. Ther. Med. 2018, 15, 3781–3790. [Google Scholar] [CrossRef]
- Huang, Y.; Du, K.L.; Guo, P.Y.; Zhao, R.M.; Wang, B.; Zhao, X.L.; Zhang, C.Q. IL-16 regulates macrophage polarization as a target gene of mir-145-3p. Mol. Immunol. 2019, 107, 1–9. [Google Scholar] [CrossRef]
- He, X.; Ji, J.; Wang, T.; Wang, M.B.; Chen, X.L. Upregulation of circulating miR-195-3p in heart failure. Cardiology 2017, 138, 107–114. [Google Scholar] [CrossRef]
- Li, F.Q.; Xu, B.; Wu, Y.J.; Yang, Z.L.; Qian, J.J. Differential microRNA expression in signet-ring cell carcinoma compared with tubular adenocarcinoma of human gastric cancer. Genet. Mol. Res. 2015, 14, 739–747. [Google Scholar] [CrossRef]
- Ma, Y.; Wu, Y.; Chen, J.; Huang, K.; Ji, B.; Chen, Z.; Wang, Q.; Ma, J.; Shen, S.; Zhang, J. miR-10a-5p promotes chondrocyte apoptosis in osteoarthritis by targeting HOXA1. Mol. Ther.-Nucleic Acids 2019, 14, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, Y.; Long, J.; Guo, W.; Ye, W. MicroRNA-195-5p inhibitor prevents the development of osteoarthritis by targeting REGγ. Mol. Med. Rep. 2019, 19, 4561–4568. [Google Scholar] [CrossRef] [PubMed]
- Lino Cardenas, C.L.; Henaoui, I.S.; Courcot, E.; Roderburg, C.; Cauffiez, C.; Aubert, S.; Copin, M.C.; Wallaert, B.; Glowacki, F.; Dewaeles, E.; et al. miR-199a-5p is upregulated during fibrogenic response to tissue injury and mediates TGFbeta-induced lung fibroblast activation by targeting caveolin-1. PLoS Genet. 2013, 9, e1003291. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, X.; Yang, M.; Ruan, W.; Wei, W.; Gu, D.; Wang, J.; Guo, X.; Guo, L.; Yuan, Y. miR-145-5p increases osteoclast numbers In vitro and aggravates bone erosion in collagen-induced arthritis by targeting osteoprotegerin. Med. Sci. Monit. 2018, 24, 5292–5300. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, H.; Xia, Y.; Yan, F.; Lu, Y. Therapeutic potential of mesenchymal cell-derived miRNA-150-5p-expressing exosomes in rheumatoid arthritis mediated by the modulation of MMP14 and VEGF. J. Immunol. 2018, 201, 2472–2482. [Google Scholar] [CrossRef]
- Mohan, A.; Singh, R.S.; Kumari, M.; Garg, D.; Upadhyay, A.; Ecelbarger, C.M.; Tripathy, S.; Tiwari, S. Urinary exosomal microRNA-451-5p is a potential early biomarker of diabetic nephropathy in rats. PLoS ONE 2016, 11, e0154055. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Wang, K.; Huang, X.; Zhao, Z.; Zhao, Z. LncRNA MALAT1 contributes to non-small cell lung cancer progression via modulating miR-200a-3p/programmed death-ligand 1 axis. Int. J. Immunopathol. Pharmacol. 2019, 33, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Sun, N.; Zeng, J.; Zeng, Y.; Fan, Y.; Feng, W.; Li, J. Differential expression of miR-672-5p and miR-146a-5p in osteoblasts in rats after steroid intervention. Gene 2016, 591, 69–73. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Ding, X.; Wang, Y.; Li, Z.; Zhao, W.; Jia, J.; Zhou, J.; Ge, J. Differential microRNA expression profiles and bioinformatics analysis between young and aging spontaneously hypertensive rats. Int. J. Mol. Med. 2018, 41, 1584–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.-F.; Zhang, J.-G.; Guo, T.-X.; Pan, X.-J. MiR-504 inhibits cell proliferation and invasion by targeting LOXL2 in non small cell lung cancer. Biomed. Pharmacother. 2018, 97, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Fearon, A.; Dahlstrom, J.E.; Twin, J.; Cook, J.; Scott, A. The Bonar score revisited: Region of evaluation significantly influences the standardized assessment of tendon degeneration. J. Sci. Med. Sport 2014, 17, 346–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dakin, S.G.; Werling, D.; Hibbert, A.; Abayasekara, D.R.E.; Young, N.J.; Smith, R.K.W.; Dudhia, J. Macrophage sub-populations and the lipoxin A4 receptor implicate active inflammation during equine tendon repair. PLoS ONE 2012, 7, e32333. [Google Scholar] [CrossRef] [Green Version]
- Dakin, S.G.; Martinez, F.O.; Yapp, C.; Wells, G.; Oppermann, U.; Dean, B.J.; Smith, R.D.; Wheway, K.; Watkins, B.; Roche, L.; et al. Inflammation activation and resolution in human tendon disease. Sci. Transl. Med. 2015, 7, 311ra173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendal, A.R.; Layton, T.; Al-Mossawi, H.; Appleton, L.; Dakin, S.; Brown, R.; Loizou, C.; Rogers, M.; Sharp, R.; Carr, A. Multi-omic single cell analysis resolves novel stromal cell populations in healthy and diseased human tendon. Sci. Rep. 2020, 10, 13939. [Google Scholar] [CrossRef]
- Hristov, M.; Erl, W.; Weber, P.C. Endothelial progenitor cells. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1185–1189. [Google Scholar] [CrossRef] [Green Version]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.H.; Al-Youha, S.; Van Agtmael, T.; Lu, Y.; Wong, J.; McGrouther, D.A.; Kadler, K.E. Tendon is covered by a basement membrane epithelium that is required for cell retention and the prevention of adhesion formation. PLoS ONE 2011, 6, e16337. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Cai, M.; Lou, L. Identification of differentially expressed genes and small molecule drugs for the treatment of tendinopathy using microarray analysis. Mol. Med. Rep. 2015, 11, 3047–3054. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Lee, F.Y.; Tarafder, S.; Kao, K.; Jun, Y.; Yang, G.; Mao, J.J. Harnessing endogenous stem/progenitor cells for tendon regeneration. J. Clin. Investig. 2015, 125, 2690–2701. [Google Scholar] [CrossRef]
- Liu, Y.; Andarawis-Puri, N.; Eppell, S.J. Method to extract minimally damaged collagen fibrils from tendon. J. Biol. Methods 2016, 3, e54. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Lui, P.P.Y.; Lee, Y.W. In vivo identity of tendon stem cells and the roles of stem cells in tendon healing. Stem. Cells Dev. 2013, 22, 3128–3140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Hua, H.; Chiu, Y.H.; Li, G.; Zhi, H.; Yu, Z.; Ren, F.; Luo, Y.; Cui, W. CD146 regulates growth factor-induced mTORC2 activity independent of the PI3K and mTORC1 pathways. Cell Rep. 2019, 29, 1311–1322.e1315. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; Wu, Z.; Duan, H.; Jiang, X.; Tu, T.; Lu, D.; Luo, Y.; Wang, P.; Song, L.; Feng, J.; et al. Impaired tumor angiogenesis and VEGF-induced pathway in endothelial CD146 knockout mice. Protein Cell 2014, 5, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Lin, Y.; Yang, D.; Shen, Y.; Yuan, M.; Zhang, Z.; Li, P.; Xia, H.; Li, L.; Luo, D.; et al. A novel anti-CD146 monoclonal antibody, AA98, inhibits angiogenesis and tumor growth. Blood 2003, 102, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Dakin, S.G.; Dudhia, J.; Smith, R.K.W. Resolving an inflammatory concept: The importance of inflammation and resolution in tendinopathy. Vet. Immunol. Immunop. 2014, 158, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.T.; Lee, C.W.; Li, M.Y.; Wang, Y.F.; Yung, P.S.; Lee, O.K. The shift in macrophages polarisation after tendon injury: A systematic review. J. Orthop. Translat. 2020, 21, 24–34. [Google Scholar] [CrossRef]
- Quirici, N.; Soligo, D.; Bossolasco, P.; Servida, F.; Lumini, C.; Deliliers, G.L. Isolation of bone marrow mesenchymal stem cells by anti-nerve growth factor receptor antibodies. Exp. Hematol. 2002, 30, 783–791. [Google Scholar] [CrossRef]
- Bühring, H.J.; Battula, V.L.; Treml, S.; Schewe, B.; Kanz, L.; Vogel, W. Novel markers for the prospective isolation of human MSC. Ann. N. Y. Acad. Sci. 2007, 1106, 262–271. [Google Scholar] [CrossRef]
- Kajikawa, Y.; Morihara, T.; Watanabe, N.; Sakamoto, H.; Matsuda, K.; Kobayashi, M.; Oshima, Y.; Yoshida, A.; Kawata, M.; Kubo, T. GFP chimeric models exhibited a biphasic pattern of mesenchymal cell invasion in tendon healing. J. Cell Physiol. 2007, 210, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E.; Mezzapelle, R. The chemokine receptor CXCR4 in cell proliferation and tissue regeneration. Front. Immunol. 2020, 11, 2109. [Google Scholar] [CrossRef]
- Sun, J.; Mou, C.; Shi, Q.; Chen, B.; Hou, X.; Zhang, W.; Li, X.; Zhuang, Y.; Shi, J.; Chen, Y.; et al. Controlled release of collagen-binding SDF-1α from the collagen scaffold promoted tendon regeneration in a rat Achilles tendon defect model. Biomaterials 2018, 162, 22–33. [Google Scholar] [CrossRef]
- Riley, G. Tendinopathy: From basic science to treatment. Nat. Clin. Pract. Rheum. 2008, 4, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, P.P.; Chan, L.S.; Lee, Y.W.; Fu, S.C.; Chan, K.M. Sustained expression of proteoglycans and collagen type III/type I ratio in a calcified tendinopathy model. Rheumatology 2010, 49, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.S.; Schizas, N.; Li, J.; Ahmed, M.; Östenson, C.G.; Salo, P.; Hewitt, C.; Hart, D.A.; Ackermann, P.W. Type 2 diabetes impairs tendon repair after injury in a rat model. J. Appl. Physiol. 2012, 113, 1784–1791. [Google Scholar] [CrossRef]
- Russo, V.; Mauro, A.; Martelli, A.; Di Giacinto, O.; Di Marcantonio, L.; Nardinocchi, D.; Berardinelli, P.; Barboni, B. Cellular and molecular maturation in fetal and adult ovine calcaneal tendons. J. Anat. 2015, 226, 126–142. [Google Scholar] [CrossRef] [Green Version]
- Howell, K.; Chien, C.; Bell, R.; Laudier, D.; Tufa, S.F.; Keene, D.R.; Andarawis-Puri, N.; Huang, A.H. Novel model of tendon regeneration reveals distinct cell mechanisms underlying regenerative and fibrotic tendon healing. Sci. Rep. 2017, 7, 45238. [Google Scholar] [CrossRef] [Green Version]
- Beredjiklian, P.K.; Favata, M.; Cartmell, J.S.; Flanagan, C.L.; Crombleholme, T.M.; Soslowsky, L.J. Regenerative versus reparative healing in tendon: A study of biomechanical and histological properties in fetal sheep. Ann. Biomed. Eng. 2003, 31, 1143–1152. [Google Scholar] [CrossRef]
- Zamboulis, D.E.; Thorpe, C.T.; Ashraf Kharaz, Y.; Birch, H.L.; Screen, H.R.C.; Clegg, P.D. Postnatal mechanical loading drives adaptation of tissues primarily through modulation of the non-collagenous matrix. eLife 2020, 9, e58075. [Google Scholar] [CrossRef]
- Gumucio, J.P.; Sugg, K.B.; Mendias, C.L. TGF-β superfamily signaling in muscle and tendon adaptation to resistance exercise. Exerc. Sport Sci. Rev. 2015, 43, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaji, D.A.; Howell, K.L.; Balic, Z.; Hubmacher, D.; Huang, A.H. Tgfβ signaling is required for tenocyte recruitment and functional neonatal tendon regeneration. eLife 2020, 9, e51779. [Google Scholar] [CrossRef]
- Patel, D.; Zamboulis, D.E.; Spiesz, E.M.; Birch, H.L.; Clegg, P.D.; Thorpe, C.T.; Screen, H.R.C. Structure-function specialisation of the interfascicular matrix in the human achilles tendon. Acta Biomater. 2021, 131, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Honnegowda, T.M.; Kumar, P.; Udupa, E.G.P.; Kumar, S.; Kumar, U.; Rao, P. Role of angiogenesis and angiogenic factors in acute and chronic wound healing. Plast. Aesthetic Res. 2015, 2, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Joshkon, A.; Heim, X.; Dubrou, C.; Bachelier, R.; Traboulsi, W.; Stalin, J.; Fayyad-Kazan, H.; Badran, B.; Foucault-Bertaud, A.; Leroyer, A.S.; et al. Role of CD146 (MCAM) in physiological and pathological angiogenesis-contribution of new antibodies for therapy. Biomedicines 2020, 8, 633. [Google Scholar] [CrossRef] [PubMed]
- Dubin, J.A.; Greenberg, D.R.; Iglinski-Benjamin, K.C.; Abrams, G.D. Effect of micro-RNA on tenocytes and tendon-related gene expression: A systematic review. J. Orthop. Res. 2018, 36, 2823–2829. [Google Scholar] [CrossRef]
- Millar, N.L.; Gilchrist, D.S.; Akbar, M.; Reilly, J.H.; Kerr, S.C.; Campbell, A.L.; Murrell, G.A.C.; Liew, F.Y.; Kurowska-Stolarska, M.; McInnes, I.B. MicroRNA29a regulates IL-33-mediated tissue remodelling in tendon disease. Nat. Commun. 2015, 6, 6774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, A.E.; Millar, N.L.; Platt, J.; Kitson, S.M.; Akbar, M.; Rech, R.; Griffin, J.; Pool, R.; Hughes, T.; McInnes, I.B.; et al. MicroRNA29a treatment improves early tendon injury. Mol. Ther. 2017, 25, 2415–2426. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The arrive guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Viera, A.J.; Garrett, J.M. Understanding interobserver agreement: The kappa statistic. Fam. Med. 2005, 37, 360–363. [Google Scholar] [PubMed]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biela, E.; Galas, J.; Lee, B.; Johnson, G.L.; Darzynkiewicz, Z.; Dobrucki, J.W. Col-F, a fluorescent probe for ex vivo confocal imaging of collagen and elastin in animal tissues. Cytom. Part A 2013, 83A, 533–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J 2011, 17, 3. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Society. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Zhou, Z.; Cong, P.; Tian, Y.; Zhu, Y. Using RNA-seq data to select reference genes for normalizing gene expression in apple roots. PLoS ONE 2017, 12, e0185288. [Google Scholar] [CrossRef] [PubMed]
- Gouin, K.; Peck, K.; Antes, T.; Johnson, J.L.; Li, C.; Vaturi, S.D.; Middleton, R.; de Couto, G.; Walravens, A.-S.; Rodriguez-Borlado, L.; et al. A comprehensive method for identification of suitable reference genes in extracellular vesicles. J. Extracell. Vesicles 2017, 6, 1347019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Liu, Y.; Du, L.; Li, J.; Jiang, X.; Zheng, G.; Qu, A.; Wang, H.; Wang, L.; Zhang, X.; et al. Identification and validation of reference genes for the detection of serum microRNAs by reverse transcription-quantitative polymerase chain reaction in patients with bladder cancer. Mol. Med. Rep. 2015, 12, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Log2 Fold Change | p-Value | FDR-adj. p-Value | |||||
|---|---|---|---|---|---|---|---|
| ID | D4 Sham v. Injured | Injured D4 v. D21 | D4 Sham v. Injured | Injured D4 v. D21 | D4 Sham vs. Injured | Injured D4 vs. D21 | Disease Associations |
| miR-365-5p | 2.21 | 0.10 | 0.00 | 0.88 | 0.08 | 1.00 | Osteoporosis [31] |
| miR-338-5p | 1.97 | 0.16 | 0.00 | 0.80 | 0.10 | 1.00 | Rheumatoid arthritis [32] |
| miR-143-3p | 1.63 | 1.10 | 0.00 | 0.02 | 0.08 | 0.79 | Rheumatoid arthritis [33] |
| miR-145-3p | 1.55 | 1.08 | 0.01 | 0.05 | 0.20 | 1.00 | Heart disease [34] |
| miR-195-3p | 1.52 | 0.00 | 0.00 | 1.00 | 0.08 | 1.00 | Heart disease [35] |
| miR-676 | 1.16 | 0.32 | 0.00 | 0.43 | 0.17 | 1.00 | Gastric cancer [36] |
| miR-10a-5p | 1.10 | 0.86 | 0.01 | 0.04 | 0.30 | 1.00 | Osteoarthritis [37] |
| miR-195-5p | 0.78 | 1.76 | 0.25 | 0.01 | 1.00 | 0.67 | Osteoarthritis [38] |
| miR-199a-5p | 0.74 | 1.29 | 0.24 | 0.04 | 1.00 | 1.00 | Lung fibrosis [39] |
| miR-145-5p | 0.71 | 1.80 | 0.36 | 0.02 | 1.00 | 0.79 | Rheumatoid arthritis [40] |
| miR-150-5p | −0.87 | −0.26 | 0.01 | 0.44 | 0.35 | 1.00 | Rheumatoid arthritis [41] |
| miR-451-5p | −1.06 | −0.37 | 0.01 | 0.39 | 0.36 | 1.00 | Diabetic neuropathy [42] |
| miR-200a-3p | −1.37 | 0.55 | 0.00 | 0.13 | 0.08 | 1.00 | Lung cancer [43] |
| miR-672-5p | −1.80 | 1.63 | 0.00 | 0.00 | 0.09 | 0.30 | Osteonecrosis [44] |
| miR-218a-5p | −2.09 | 0.72 | 0.00 | 0.24 | 0.08 | 1.00 | Hypertension [45] |
| miR-504 | −3.38 | 1.11 | 0.00 | 0.16 | 0.09 | 1.00 | Lung cancer [46] |
| ID | Gene ID | Log2 Fold Change: Sham vs. Injured (Day 4) |
|---|---|---|
| miR-365-5p | MIMAT0017184 | 1.49 |
| miR-338-5p | MIMAT0004646 | 1.23 |
| miR-143-3p | MIMAT0000849 | 2.79 |
| miR-195-3p | MIMAT0017149 | 0.37 |
| miR-672-5p | MIMAT0005327 | −4.06 |
| miR-218a-5p | MIMAT0000888_1 | −2.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marr, N.; Meeson, R.; Kelly, E.F.; Fang, Y.; Peffers, M.J.; Pitsillides, A.A.; Dudhia, J.; Thorpe, C.T. CD146 Delineates an Interfascicular Cell Sub-Population in Tendon That Is Recruited during Injury through Its Ligand Laminin-α4. Int. J. Mol. Sci. 2021, 22, 9729. https://doi.org/10.3390/ijms22189729
Marr N, Meeson R, Kelly EF, Fang Y, Peffers MJ, Pitsillides AA, Dudhia J, Thorpe CT. CD146 Delineates an Interfascicular Cell Sub-Population in Tendon That Is Recruited during Injury through Its Ligand Laminin-α4. International Journal of Molecular Sciences. 2021; 22(18):9729. https://doi.org/10.3390/ijms22189729
Chicago/Turabian StyleMarr, Neil, Richard Meeson, Elizabeth F. Kelly, Yongxiang Fang, Mandy J. Peffers, Andrew A. Pitsillides, Jayesh Dudhia, and Chavaunne T. Thorpe. 2021. "CD146 Delineates an Interfascicular Cell Sub-Population in Tendon That Is Recruited during Injury through Its Ligand Laminin-α4" International Journal of Molecular Sciences 22, no. 18: 9729. https://doi.org/10.3390/ijms22189729